Soybean meal depletion impaired growth in pigs despite restoration of all essential amino acids and non-essential amino acid nitrogen
By R. Dean Boyd, Mike Johnston, James Usry and Richard E. Austic
This study of low protein diets for pigs was reported 25 years ago by Johnston and coworkers (1999), in anticipation that the library of free amino acids for commercial use would expand during the next decade. Two amino acids were in use at that time (DL-methionine, L-lysine) with threonine expected to be available within a few years. The driving force for this expansion was to reduce dependency on SBM as the primary source of amino acids, while reducing diet cost. A secondary factor was to reduce nitrogen excretion.Attempts to reduce dietary crude protein (CP) were successful to a point, but the extent to which SBM could be replaced by corn and synthetic amino acids was less than expected. Eventually, growth was impaired and feed cost of gain increased. The reason for this was unclear, but the order of the 4th and 5th most limiting amino acids for a corn – soybean meal diet (C-S) was uncertain. This was complicated by the absence of reliable amino acid digestibility estimates for ingredients. Attempts to reduce diet CP level by 4-5 percentage points (e.g. 16% to 12%) were met with failure to maintain growth, which led to the proposition that non-essential amino acids (NEAA) may have become deficient.
These obstacles caused our team to take a different approach to the study of low CP diets in order to determine if feeding a diet with little intact protein (beyond corn) was even possible. The most important question was whether there was a level of SBM (or intact protein) displacement that would result in impaired growth, even with complete restoration of all essential amino acids (EAA) and NEAA.
Our mandate, as nutritionists for a genetic improvement firm, was to specify diets that did not (1) constrain genetic expression for growth rate, feed conversion efficiency (FCE), protein deposition or carcass lean, and that (2) minimized feed cost and environmental impact, while doing so.
What was learned?
This study involved 48 pigs in the 118 to 188 lb phase of growth and suggested that there was a point in SBM or intact protein displacement where growth would start to become impaired. This is often expressed by inefficient FCE, while growth rate may or may not be reduced, depending on how feed intake is affected. FCE was impaired by ‘extreme’ SBM displacement, which coincided with pigs becoming fatter. This occurred even though low CP diets were made equivalent to the C-S control diet in all 10 EAA and NEAA nitrogen. Subtle changes in diet electrolyte balance (DEB), K and choline content were controlled.
This study was the foundation to subsequent studies by the Pig Improvement Co. (PIC), to make certain that genetic expression was not constrained by diet as the free amino acids library expanded. Results of this study suggested that, at some level of free amino acid use, something other than amino acids would become first limiting to growth (e.g. SBM or intact protein). Although our results were mystifying, we concluded that there was room to expand the free amino acids library, but unrestricted use would not be possible without emergence of new knowledge. Younger swine nutritionists assume that free amino acids can be used without restraint, provided that the ideal amino acid pattern is met. Nothing could be further from the truth.
Purpose and logic for this study
This study was designed to determine if the impaired growth that occurs with extensive SBM (or CP) removal could be prevented by restoring all EAA and NEAA nitrogen (N) that was lost in the substitution of corn for SBM. Prior to this time, this approach has been limited to studies with laboratory rats (Dr. N. Benevenga, pers. comm.) because complete replacement of all amino acids was expensive. Low CP diet studies with pigs typically involved replacement of those EAA that were considered to be deficient based on an Ideal amino acid pattern for the time (Baker, 1997).
Growth measures included rate and efficiency of weight gain, but weight gained was defined by its composition (protein, lipid, water, ash) in order to clarify the effect of diet on the nature of growth. For example, if weight gain for low CP diets was equal to the control diet and composition of that gain retained the original balance in lipid and protein content, then growth was not impaired. Conversely, if weight gain on the low CP diet was equal to the control diet because of an increase in feed intake, then composition of that gain (protein, lipid) may have been altered, which is normally reflected by FCE response.
Methods overview
This study was conducted at the PIC USA research center in Franklin KY. It involved a growth assay from 118 to 188 lbs with harvest immediately afterward to characterize the composition of weight gained. Forty-eight female pigs (PIC terminal genetics) were allotted to 1 of 6 dietary treatments (8 EU/diet) and penned individually (12 ft2/pig net of feeder space) with ad libitum access to diet and water. Pooled semen from 6 sires was used to minimize paternal variation for progeny growth. Candidate pigs were selected based on weight gain/day of age (<8% range) and allocated to diets balanced for litter origin. Room temperature was maintained thermo-neutral with light pattern set to a 16-h light, 8-h dark cycle for the 31 d growth assay.
Initial whole-body composition was established using 9 contemporary pigs with feed being withdrawn 14-16 h prior to harvest (fasted body weight, 103 + 5.8 SEM). Harvest and chemical analysis was conducted by the University of Illinois Meat Lab (Dr. F. McKeith). The whole-body, including head, organs and gastrointestinal tract (minus residual digesta), was ground and analyzed for water, protein, lipid and ash content according to AOAC procedures (1980). Of the 48 pigs used for the growth assay, 36 of them (6 pigs/diet) were harvested to estimate body composition at the conclusion of study (approx. weight constant basis). Lipid and protein accretion rates were calculated by standard balance procedures.
This study was anchored on a (1) nutritionally adequate positive control diet (PC, 17% CP) to establish the expressed genetic capacity for growth; (2) negative control diet (NC, 15% CP) formulated to be slightly deficient in lysine so that growth rate and FCE would be inferior to the PC diet. This was the reference diet to which pigs fed the low CP diets were compared; (3) serial reductions in SBM content (13, 11% CP). Measures of growth included: growth rate, FCE, daily feed intake, protein and lipid deposition rates (Pd, Ld).
Design for diet comparison
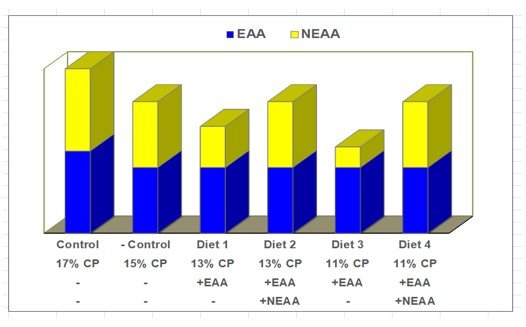
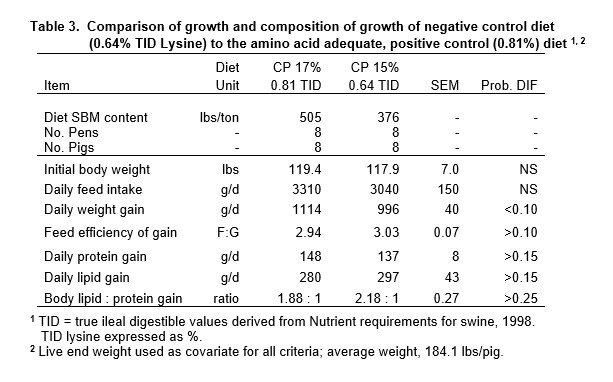
The framework for all dietary treatments is shown in Figure 1. The PC diet was formulated to 0.81% true ileal digestible lysine (TID) using digestibility coefficients from the Nutrient Requirements of Swine (NRC, 1998) for corn and SBM. This lysine specification was derived from dose-response studies, for the genetic lines of pigs used. It represents the maximum response (Rmax) to TID lysine. The ideal pattern for other EAA to lysine met or exceeded 1998 NRC specifications (TID basis). The reference diet was formulated to 0.64% TID lysine, a level proven from the lysine growth assay curve to support sub-maximum growth (95% of Rmax). This met guidelines for a sensitive reference point diet (Baker, 1986).
The decline in CP from the PC (17%) to the NC reference (15%) was achieved by displacing SBM with ground corn. Further reductions in SBM content were made to create 13% and 11% CP diets, but TID lysine (0.64%) and all EAA in these diets were restored to the level of the reference by adding all 10 EAA in exchange for corn. The conditionally essential amino acids (CEAA), L-cysteine and L-tyrosine, were added to ensure that methionine and phenylalanine were not used to synthesize them (NRC, 1998).
A second pair of low CP diets (13, 11% CP) were made equivalent to the previous pair, with the exception that 2 NEAA (L-aspartic and L-glutamic acids) and 1 CEAA (L-glycine) were added to the level of NEAA nitrogen in the reference diet. Aspartic acid, glutamic acid and glycine were provided as substrates for the synthesis of NEAA, in case 1 or more of them became deficient by diet CP reduction.
This design allowed us to determine if serial reductions in SBM, to create 13 and 11% CP diets, could support growth equal to the 15% CP reference diet, provided that all EAA were restored (first diet pair) or if NEAA N sources were also needed to maintain growth equivalent to the reference.
Diet composition and nutrient specifications
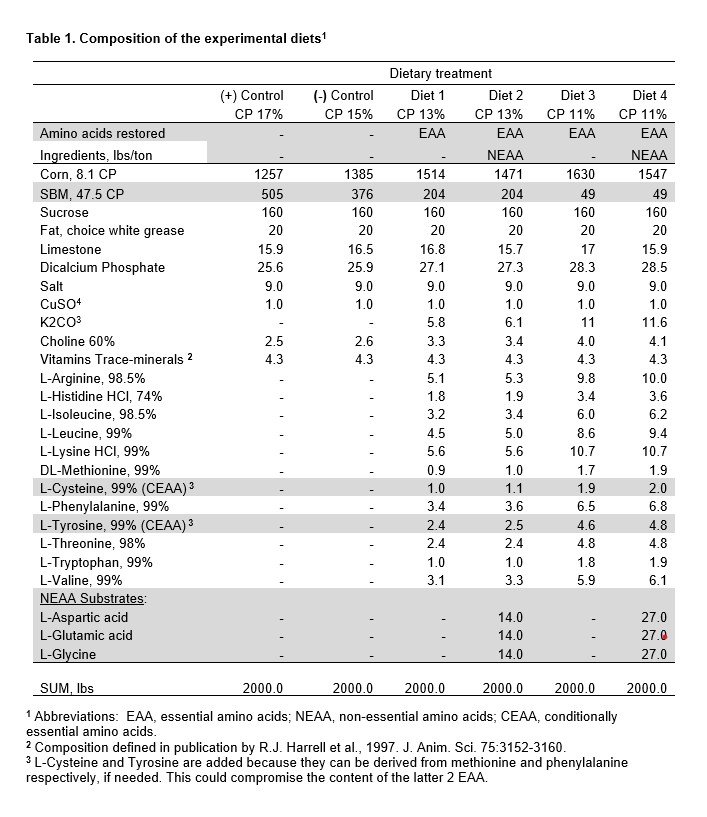
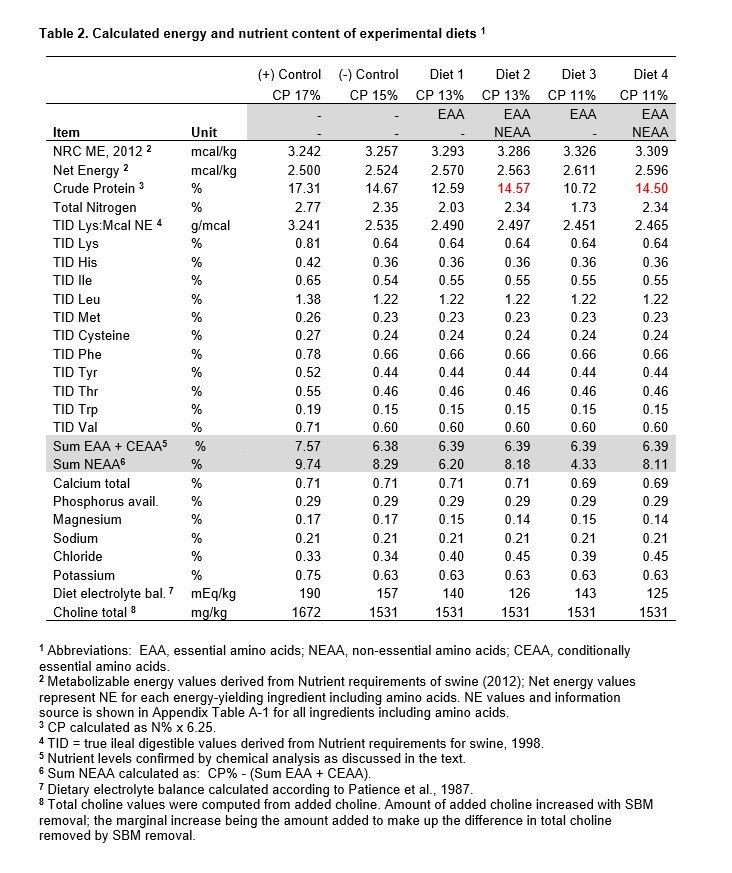
Diet composition is shown in Table 1. Serial SBM displacement resulted in extreme dietary CP decline (17, 15, 13, 11% CP). This 35% decline in CP was accomplished by a 90% reduction in SBM content; declining from a high of 505 lbs/ton in the PC diet to 49 lbs/ton for 11% CP diets. Diet equivalence for all other nutrients was achieved. Choline was added to offset the decline in total choline by SBM removal. K2CO3 was added to restore the loss of K from SBM and to maintain dietary electrolyte balance (DEB), above a level that has proven safe for feed intake (Patience et al., 1987). Electrolyte balance countered Cl- influx from commercial sources of lysine and histidine.
Since net energy (NE) estimates were lacking for the spectrum of amino acids used and the NE value for SBM (NRC, 1998) was considered to be inaccurate, isocaloric diets could not be formulated. For this reason, dietary fat was held constant for diets. Theoretical estimates suggested that 13% CP and 11% CP diets may have higher NE values than the reference diet, by up to 3%. This was viewed as a complication for interpretation, but not a fatal flaw.
The possibility that extensive crystalline amino acid use could harm diet palatability was countered by adding sucrose to all diets. Analytical values for corn and SBM amino acid levels were provided by the Continental Grain feed manufacturing plant, in Hopkinsville, KY (now Hubbard Feeds). Diets were mixed at the feed mill by Mike Johnston, with staff assistance. Johnston also prepared EAA and NEAA premixes. The calculated nutrient content for diets is summarized in Table 2. Amino acid levels were chemically confirmed on extensively sampled diets (< 6% variance with few exceptions).
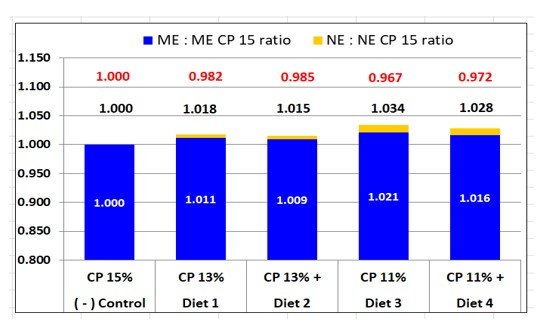
Credible NE estimates for SBM recently became available from the Hans Stein lab (Lee et al., 2021 and 2022), that agreed with growth assay results conducted under academic conditions (Boyd and Gaines, 2023). Net energy estimates for the amino acids (15) that we used also became available (Rostagno et al., 2017). We computed dietary NE values from all energy-yielding ingredients using these values and estimates provided by the NRC (2012) for ground corn, sucrose and fat (choice white grease). Results of this belated diet calculation are shown in Table 2. Relative diet differences in NE (and ME) are shown in Figure 2. Ingredient NE values and references are provided in Appendix Table A-1.
Calculations, statistical analysis and planned comparisons
Composition of weight gain data was evaluated for credibility by comparing the sum of water, lipid, crude protein (N x 6.25) and ash accretion estimates to whole-body weight gain for the growth period (n = 36 pigs grown, harvested). We determined that the percentage of body weight gain that was accounted for by the 4 chemical constituents was within expectations and were relatively consistent across diets. They averaged 97.1% of whole-body gain (SD + 2.5%), with residual chemical matter being 2.9% by difference (e.g. glycogen, nucleotides, CP factor error). This suggested that our estimates for lipid and protein accretion rates were credible.
Data were analyzed using SAS PROC GLM procedure, with 5 planned comparisons: PC vs NC; Linear and non-linear response, diets NC, 1, 3; NEAA importance, diets 1, 3 vs 2, 4; low protein diet comparison, 13% CP vs 11% CP, provided that there was no interaction (see Figure 1 for diet identity).
Reference diet proven sensitive control for low protein diets
The reference diet (15% CP) was designed to be slightly deficient in lysine in order to eliminate excess lysine that could prevent immediate response if pigs fed low CP test diets began to show impaired growth. The reference proved to be deficient in 1 or more EAA, since it was unable to support the growth rate achieved by pigs fed the PC diet (Table 3, P<0.10). The FCE response tended to be inferior, which coincided with a fatter composition of gain (Ld : Pd ratio increased by 15.9%).
Extreme SBM reduction impaired growth despite restoring all EAA
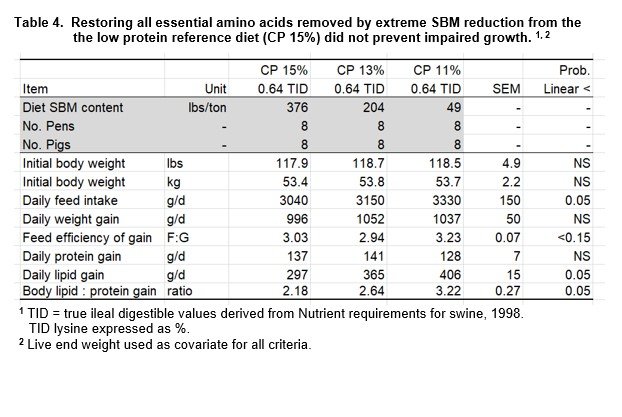
The SBM content of the 15% CP reference diet (376 lbs/ton) was reduced to 204 and 49 lbs/ton to create 13% and 11% CP diets, respectively (Table 1). Since every EAA that was reduced by removing SBM was restored to the reference level, diets differed in the content of SBM, CP and NEAA (Table 2).
Pigs fed low CP diets (13%, 11%) with EAA restored exhibited similar growth rate to pigs fed the reference diet, but composition of gain was changed (Table 4). Daily feed intake increased in a linear manner (P<0.01) for pigs fed reduced SBM diets and pigs became fatter. This increase in feed intake coincided with a linear increase in lipid deposition rate and the lipid to protein ratio of gain (P<0.05). The balance increased from 2.18 to 2.64 to 3.22 g lipid/g protein deposited (+21, +48%) for 15%, 13% and 11% CP diets, respectively.
Since restoration of EAA and CEAA (cysteine, tyrosine) did not prevent impaired growth (composition), the next step was to determine if NEAA had become deficient with dietary CP reduction (17, 15, 13, 11% CP). It seemed inevitable that extreme CP reduction would lead to insufficient substrate for NEAA synthesis.
Extreme SBM reduction impaired growth despite restoring EAA and NEAA
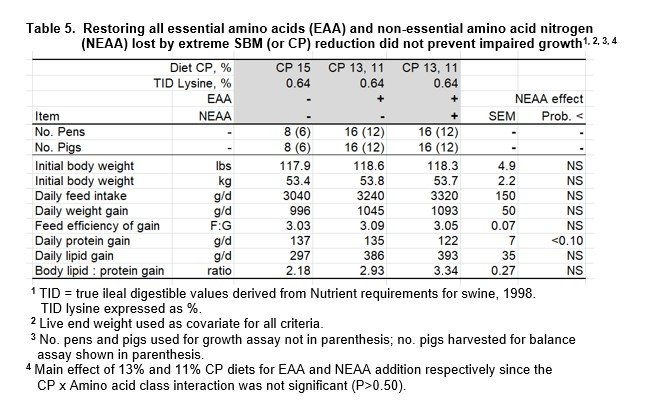
This comparison involved 2 low CP diet pairs (13%, 11% CP); one set having every EAA restored, as discussed above, and a second pair having both EAA and NEAA N restored (Table 2). Since the interaction of protein level (13, 11%) by amino acid class (EAA, EAA+NEAA) was not significant (P>0.50), the main effect of amino acid type is presented (Table 5). Although the extensive reduction of SBM (or CP) content did not alter daily growth rate, feed intake and composition of that gain was adversely affected even though all EAA and NEAA N was restored (Linear response, P<0.05).
The premise that NEAA nitrogen was equivalent to the reference diet is based on the assumption that glycine, aspartic acid and glutamic acid were appropriate substrates for synthesis of 5 other NEAA (Table 1). We emphasize that calculated levels of amino acids were chemically confirmed (< 6.0% deviance).
Big picture of diet effects on rate and composition of growth
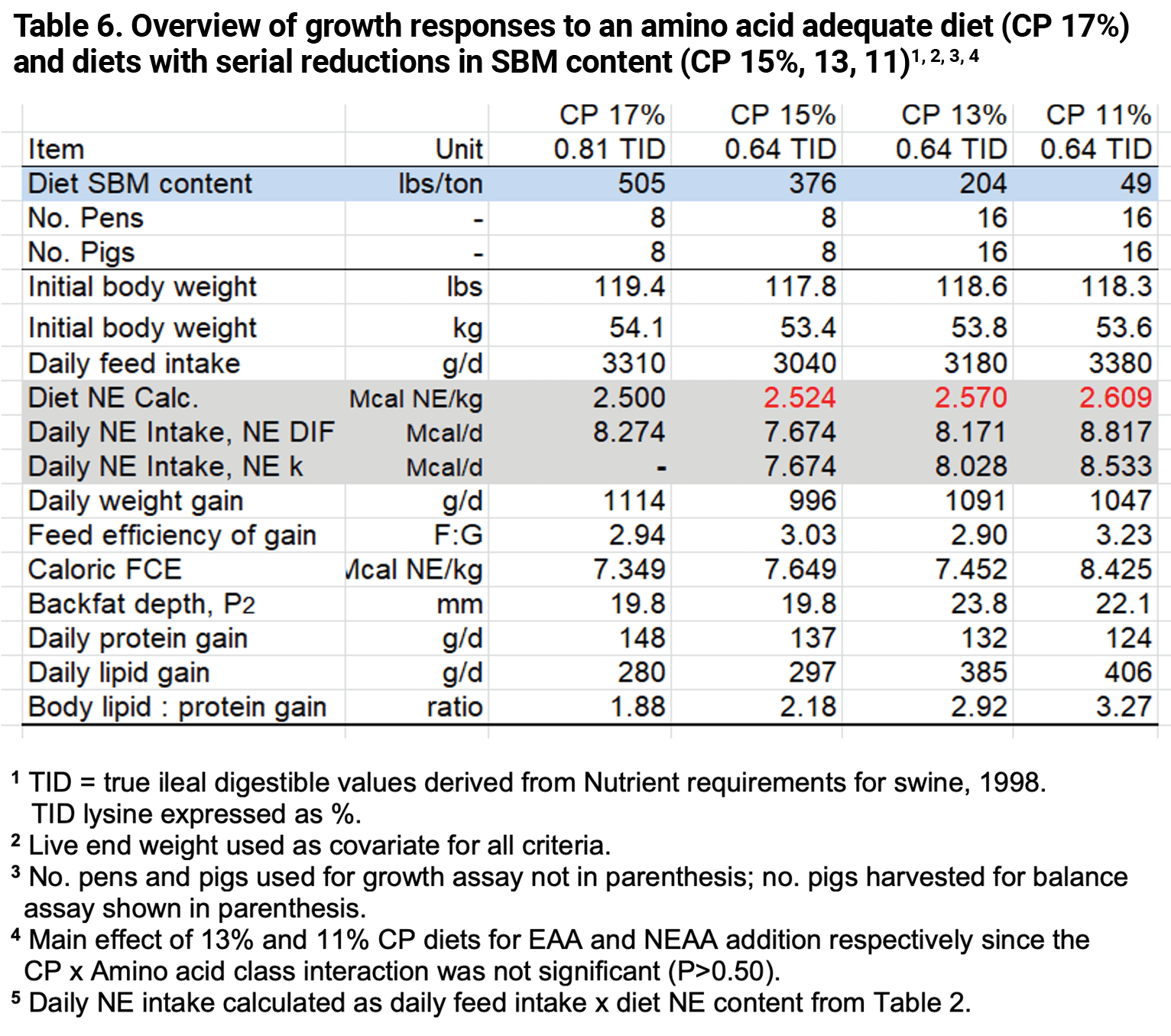
An overall summary of how experimental diets affected growth and composition of gain is presented in Table 6. Since there were no differences in response to the restoration of EAA as compared to EAA + NEAA results were pooled within each low CP diet pair (13, 11%) to improve estimated effects.
The linear increase in daily feed intake as dietary CP decreased (15, 13, 11%) suggests that intake was responding in a coordinated manner to some deficit or imbalance (Simpson and Raubenheimer, 2012) that increased as the dietary CP level declined. Daily feed intake (lbs, calories) and lipid content of weight gained (Ld and Ld : Pd ratio) showed a progression of impaired growth as diet CP declined from 15% CP to the 11% CP diet. The result was that weight gain was the ‘fattest’ for pigs fed the 11% CP diet.
The basis for the impaired growth is unclear since crystalline EAA and NEAA were equivalent for 15, 13, and 11% CP diets and since dietary nuances that occurred with SBM depletion were standardized. The most glaring difference between low CP diets was SBM content (intact protein). The 13% CP diet had 46% less SBM than the reference diet (204 vs 376 lbs/ton), while 11% CP diets had 90% less SBM (49 vs 376).
It is conceivable that progressive growth impairment is more related to intact protein (or source) than CP level. The reference diet (15% CP) had the same level of calculated CP (N% x 6.25) as the 13 and 11% CP diets containing NEAA substrates (14.7, 14.6, 14.5% CP).
How diets compared for NE content based on current NE values
Dietary NE estimates were belatedly calculated using recent ingredient estimates (Table 2). Differences in diet NE were relatively small with a subtle increase as intact protein declined (15% CP to 11% CP, Figure 2). NE for diets 1 and 2 (13% CP) were greater than for the reference diet (1.8, 1.5% respectively). NE variance for diets 3 and 4 (11% CP) increased further (3.4, 2.8% respectively). This led to a slight decline in a key determinant of growth, FCE and composition of gain (g TID Lys:Mcal NE); being 98.2 and 98.5% of the reference for 13% CP diets and 96.7 and 97.2% for 11% CP diets.
Reducing dietary CP from 15% to 11% exerted a slight NE bias that favored the 11% CP diet, so the question became whether this magnitude of NE variance could have altered the composition of gain in a measurable way. We believe that the 3.1% disadvantage in lysine to NE ratio for 11% CP diets was too small to elicit a 37% increase in Ld rate (406 vs 297 g/d) and a 50% increase in the Ld to Pd balance of gain (3.27 vs 2.18), compared to the reference diet (Table 6). By comparison, a 22% reduction in the lysine to NE ratio for the 15% CP reference diet compared to the PC diet resulted in only a 16% increase in the Ld to Pd ratio for gain (Tables 2, 6).
We conclude that something other than a slight caloric bias in the lysine to NE ratio (-1.7 and -3.1% for 13 and 11% CP diets) caused growth to be significantly impaired.
How our results compare with 21st century low protein diet research?
Attempts to reduce dietary CP have been successful for a 2-3 percentage unit decline (e.g. 17 to 15%). Our attempt to extensively reduce protein level (27% from reference) without impairing growth was not successful. Complete restoration of EAA rendered ideal amino acid considerations moot; CEAA provision and NEAA substrate nitrogen was addressed.
Our findings were compared with more recent low protein diet studies that involved practical diets and the expanded library of commercial sources of free amino acids available for test diet formulation.
Nebraska Lewis – Miller Labs
Comparisons were limited to 3 papers with extensive diet protein reduction (> 25%). Two well-designed studies involving C-S diets and pigs in the early growing stage were conducted by the Nebraska lab group. Pigs were high lean genetics, considered high health and housed in individual pens. Their academic setting may be important to eliminate the possibility of confounding by extra-nutrient factors in SBM; factors that may be expressed in response to stressors in the commercial environment. This distinction was critical to distinguish between classic energy substrate NE estimates for SBM and the extra-nutrient increase in total diet NE use for growth; an effect we ascribe to complementary factors that may improve physiological conditions to support growth (Boyd and Gaines, 2023; Petry et al., 2024).
In the first study (Gomez et al., 1999), SBM was reduced 50% (11.7% CP) compared to the reference diet (15.9% CP). Diets were made equivalent for TID levels of lysine, methionine, threonine and tryptophan. Over the 72 to 130 lbs phase of growth, pigs fed the low CP diet exhibited impaired and fatter growth, based on inferior growth rate, FCE and protein deposition rate. Although lipid deposition rate was not changed (P>0.05), whole-body Ld : Pd balance was numerically greater (+23%).
The second study (Figueroa et al., 2001), involved 3 additional amino acids (valine, isoleucine, histidine) to address possible deficits in the 7 most limiting EAA. SBM was reduced by 64 to 67% in 3 experiments. The average starting weight for pigs ranged from 43 to 48 lbs (5 week study) across the experiments. Pigs were housed in individual pens. A 16% CP reference diet was compared to 2 low CP diets (12%, 11%). Pigs fed the 11% CP diet exhibited impaired growth, even though isoleucine and valine were added (Exp. 1). Pigs fed the 11% CP with combinations of valine, isoleucine and histidine, added to match the 16% reference diet, produced conflicting results. In Exp. 2, the impaired growth was not resolved, whereas in Exp. 3 growth appeared to be maintained. The difference in results for experiments 2 and 3 may be due to having only 6 pigs per diet.
INRA Le Floc’h – van Milgen Labs
The study by Gloaguen and co-workers (2014) provided an excellent comparison of results. The INRA studies (St-Gilles, France) involved extreme reductions in dietary protein and SBM, being displaced by cereal grains and amino acids. Wheat and barley were the primary grains with corn content being minor. Use of grains other than corn is noteworthy because the U.S. are founded on corn with by-product grain products. Canadian diets tend to be more diverse in cereal grains used and these differ in type and content of functional compounds (Gani et al., 2012; Singh and Sharma, 2017). INRA studies were conducted in an academic setting with pigs in the late nursery stage (22-44 lbs) and individually housed.
In Exp. 1, the reference diet had 19.7% CP with a corresponding lysine level below the requirement for the genetic product (1.15% SID lysine). Test diets contained 19.7, 16.8, 14.0 and 12.7% CP; each being fortified with up to 10 EAA to meet the lysine specification and ideal amino acid standard (NRC 2012). Reduction of CP content, from 19.7% to 16.5%, had no effect on nitrogen (N) retention, growth rate or FCE; however, the decline to 14.0% and 12.7% CP yielded inferior results. Their conclusion was appropriate, but FCE means for low CP diets were slightly charitable since diets were not isocaloric (NE basis). Compared to the reference diet, NE content increased (+3.0, 6.3, 6.7%) as CP level declined.
Exp. 2 was conducted to determine if the decline in dietary CP could be extended by including CEAA amino acids (arginine, glycine, glutamine, proline) as substrate for NEAA synthesis. Diet CP was reduced to the point of complete SBM removal by wheat and barley and free amino acids. Six dietary CP levels were used that contained SBM (17.6, 15.6, 13.5, 11.8% CP) or were devoid of SBM (14.0, 13.0% CP). The authors concluded that decreasing diet CP content from 17.6 to 13.5% had no effect on performance. Feeding a diet formulated without SBM (13.0% CP), being replaced by wheat (50%), barley (28.4%), all 10 EAA and the CEAA resulted in inferior growth and FCE. Increasing diet CP from 13.0 to 14.0% by nearly doubling the glutamine supplement restored growth rate and FCE to equal the reference. They concluded that SBM can be completely replaced by wheat – barley cereals and free amino acids to a limit of about 13.5% CP, without compromising growth.
The authors were able to reduce CP, but the decline was limited to about 3.6 percentage units (Exp. 2) given SBM replacement with ‘high’ CP grains. Diets for exp. 2 were not isocaloric. The 14% CP diet had a 5.1% advantage in NE compared to the reference diet. Taking this into account, the 14% CP diet required 6.9% more NE per kg gain (18.57 MJ/kg gain vs 17.37); caloric FCE was an improved basis for evaluation since diets were not isocaloric.
This is not a criticism, but a clarification that must be taken into account when deciding whether a low CP amino acid supplemented diet is successful in preserving growth equal to the reference. It is not absolutely clear to us whether NE values were included for some or all the amino acids used in their studies. If these were not estimated, then NE inequality among diets would increase.
Nutrient management and intact protein perspective
The value of N (and phosphorus) in pig manure is viewed differently in North America (NA) than for European countries, in general. For many in NA, manure from pig farms is safely stored and injected into crop land in an environmentally responsible manner. Organically derived N (and phosphorus) sources are used to reduce dependence on more expensive inorganic sources of N fertilizer.
There is another consideration. Intact protein from certain sources may benefit growth and FCE when pigs in the commercial setting encounter immune stress (Boyd et al., 2023). Nutritionists tend to view intact protein as only an amino acid source, but digestion can also yield small peptides that become bioactive upon release from protein. This is true for whey protein (Moughan and Rutherford-Markwick, 2013), which has recently and repeatedly been shown to improve nursery pig viability (Dr. Aaron Gaines, pers. com.). Nature has encrypted specialized chains of amino acids in SBM that are capable of improving intestinal function and (or) systemic health (Petry et al., 2024).
Low protein – SBM displaced diets require field validation
Low protein diets with a reduction greater than 2-3 percentage units need to be validated in the commercial setting to probe for unintended consequences. This is especially true when SBM (perhaps other legume seeds) are displaced because its content of functional compounds is high. Reducing diet SBM content to near zero will probably have unintended consequences, that may have little to do with amino acid nutrition. Contributions of SBM as an ingredient involves nutrient and non-nutrient health improving factors shown to counter certain disease in humans and potentially for pigs (Petry et al., 2024).
Validating concepts in the commercial setting is important because it presents a dimension of stress that is not present in the academic environment; stressors that are a function of population dynamics (pathogen, physical). SBM has been shown to elicit benefits that are more diverse than can be ascribed to nutrients alone or measured by growth studies (improved diet NE use, viability, medical treatment; Smith et al., 2020; Cemin et al., 2021; Boyd and Gaines, 2023; Petry et al., 2024).
Validation studies, in the commercial setting, need to cover an extended time (e.g. wean to market) so the life-time consequence of accruing benefits can be manifested. Measures need to be extensive (growth, FCE, medical, viability measures) and growth measures need to be related to the ultimate value outcomes of full-value pigs sold, carcass growth and, where possible, saleable meat per carcass.
Why extreme low protein diets impaired growth – nutrition meets physiology
The problem of extreme low protein diets and impaired growth remains unresolved. At some point in dietary SBM (or intact protein) displacement, by free amino acids and grain, growth fails to be maintained. We observed that pigs fed low CP diets had inferior FCE and fatter weight gain, despite restoration of all EAA and NEAA nitrogen, equivalent to the reference. Clearly, protein content of swine diets can be reduced and replaced by amino acids, but something other than amino acids eventually becomes first limiting to growth.
Principles of amino acid nutrition and of functional compounds in protein sources are better understood today than when we conducted this study 25 years ago (Johnston et al., 1999). For this reason, our perspective on possible limitations is better. Amino acid considerations of balance and deficiency were controlled by design. Although differential uptake (timing) of amino acids from free amino acids as compared to intact protein is a worthy consideration (e.g. efficiency of use question), we anticipate that functional compounds in SBM is the reason that it cannot be completely eliminated without impairing growth.
Complementary physiological factors
To be specific, functional compounds in SBM may improve metabolism outcomes (e.g. ROS- removal) and (or) ‘dampen’ inflammation that diverts energy from growth (Demas et al., 2011). Cereal grains (wheat, barley, corn) also contain functional components that may or may not contribute enough alone, but in combination with SBM they may impact growth in a way that amino acids and some grain by-products cannot. This topic is complex, but a targeted translational review of this subject was written to help animal nutritionists become more familiar with this area (Petry et al., 2024).
These compounds might be viewed as complementary physiological factors. This term is applied in the context that when they are present with another factor (e.g., nutrients), the combination produces an effect that is qualitatively distinct from the separate effects of nutrient intake alone. Their effect may be to moderate immune response or to remove harmful products of metabolism for an improved overall outcome. This phenomenon may be more demonstrable under the stress of the commercial environment.Refining the growth influences framework
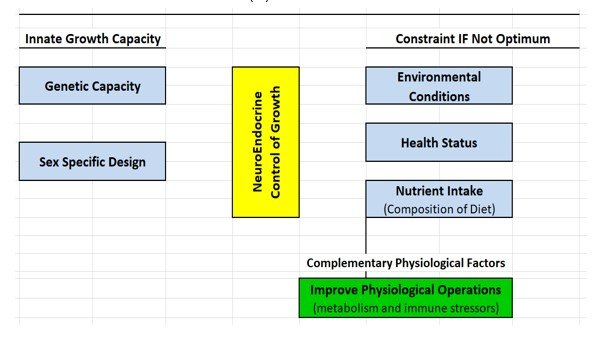
The possibility that functional compounds may account for the eventual failure of low protein diets to maintain growth was raised previously (Lewis, 2001). In light of the results of this low protein study and those of others conducting low protein diet research, a physiological complement was inserted into the framework of factors that influence growth (Figure 3). In this framework, the neuro-endocrine system (N-E) is the centerpiece that ultimately governs the amount and type of growth expressed. This sophisticated control matches the genetic capacity for growth (driver) with inputs that accommodate or constrain, but do not drive growth (Boyd and Bauman, 1989; Decuypere and Buyse, 2005). N-E control mechanisms also orchestrate changes in food intake in response to the environment, nutrient deficit or imbalance (NRC, 1987)
Nutrients accommodate N-E coordinated tissue growth (e.g. IGF-1 drives muscle deposition), but they cannot override limits imposed by the environment or health stress. This principle was illustrated in a study we conducted with pigs that encountered respiratory immune stress (Boyd et al., 2023). Amino acid level was important to growth, but secondary to dietary SBM level. The important concept incorporated into Figure 3 is that a dietary mix of ingredients provides nutrients and functional components from certain ingredients that may be concentrated enough to improve metabolic outcomes and (or) dampen inflammatory insult.
Conclusion
Dietary protein can be reduced and replaced by amino acids and grain to a point (e.g. 2.5 to 3% CP).
Severe depletion of SBM eventually impaired growth (esp. FCE) even though all essential amino acids and non-essential amino acid nitrogen was restored to match the reference diet.
There is a point in SBM depletion where something other than amino acids appears to become first limiting to normal growth.
Functional molecules may be complementary to optimum metabolic and immune function in the commercial environment.
Not all alternative protein sources are equal to SBM and other legumes in functional compounds content. Some impair feed intake (e.g., DDGS), which is a problem for summer carcass weight targets.
Although we are open to the possibility that some combination of free amino acids might eliminate impaired growth that occurs with extreme reductions in SBM, we believe that the study of SBM as a source of protein and functional compounds will be more immediately fruitful. We are less inclined to consider minimum intact protein per se than the broader benefits of a protein ingredient for a commercial environment.
Reference Information
Johnston, M.E., J.L. Usry and R.D. Boyd. 1999. Effect of feeding low protein diets on performance of growing pigs. J. Anim. Sci. 77 (Suppl.1):69.
Baker, D.H. 1997. Ideal amino acid profiles for swine and poultry and their application in feed formulation. Biokyowa Tech. Rev. 9. Biokyowa Inc.
AOAC. 1980. Official Methods of Analysis. 13th ed. Assoc. Official Anal. Chem., Washington, DC
National Research Council (NRC). 1998. Nutrient requirements of swine 10th revised edition. National Academy Press, Washington, DC, USA.
Baker, D.H. 1986. Problems and pitfalls in animal experiments designed to establish dietary requirements for essential nutrients. J. Nutr. 116:2339-2349.
Patience, J. F., R. E. Austic, and R. D. Boyd. 1987. Effect of dietary electrolyte balance on growth and acid-base status in swine. J. Anim. Sci. 64:457–466.
Lee, S.A., D.A. Rodriquez and H.H. Stein. 2021. Determination of the net energy in soybean meal fed to pigs using indirect calorimetry. Report to United Soybean Board, USB Res. Proj. 2030-353-0515-G
Lee, S.A., D.A. Rodriguez and H.H. Stein. 2022. Net energy in U.S. soybean meal fed to group-housed growing pigs is greater than calculated book values. 15th Int. Symp. Diges. Physio. Pigs. Anim., Sci. proc. 13 (Issue 2):178.
Boyd, R.D. and A.M. Gaines. 2023. Soybean meal NE value for growing pigs is greater in commercial environments. Feedstuffs, August digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-august-2023/page/2/1
Rostagno, H.S. et al., 2017. Brazilian tables for poultry and swine: Composition of feedstuff and nutritional requirements. 4th ed. Dep. Zootecnia, Univ. Fed. Vicosa MG
National Research Council (NRC). 2012. Nutrient requirements of swine 11th revised edition. National Academy Press, Washington, DC, USA. https://doi.org/10.17226/1329
Simpson S.J. and D. Raubenheimer. 2012. The nature of nutrition: A unifying framework from animal adaptation to human obesity. Princeton University Press.
Gomez, S., P.S. Miller, A. Lewis and H.Y. Chen. 1999. Growth and carcass responses of barrows fed a corn-soybean meal diet or low-protein amino acid-supplemented diets at two feeding levels. Nebraska Sw. Rpts 131:35-39. https://digitalcommons.unl.edu/coopext_swine/131
Figueroa, J. L., A. J. Lewis, P. S. Miller, R. L. Fischer, and R. M. Diedrichsen. 2003. Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. J. Anim. Sci. 81:1529–1537
Gloaguen, M., N. Le Floc’h, E. Corrent, Y. Primot and J. van Milgen. 2014. The use of free amino acids allows formulating very low crude protein diets for piglets. J. Anim. Sci. 92:637-644. https://doi.org/10.2527/jas2012-6514
Gani, A., S.M. Wani, F.A. Masood and G. Hameed. 2012. Whole-grain cereal bioactive compounds and their health benefits: A review. J. Food. Process Technol. 3:3. http://dx.doi.org/10.4172/2157-7110.1000146
Singh, A. and S. Sharma. 2017. Bioactive components and functional properties of biologically activated cereal grains: A bibliographic review. Critical Reviews in Food Sci. Nutr. 57 (14): 3051-3071. https://doi.org/10.1080/10408398.2015.1085828
Petry, A., B. Bowen, L. Weaver and R. Dean Boyd. 2024. Functional compounds in soybean mean: implications for pig health and physiology. Feedstuffs, February digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-february-2024/page/5/1
Boyd, R.D., M.E. Johnston, J. Usry, P. Yeske and A. Gaines. 2023. Soybean meal mitigates respiratory disease-impaired growth in pigs. Feedstuffs, October digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-october-2023/ page/2/1.
Moughan, P.J. and K. Rutherford-Markwick. 2013. Food bioactive proteins and peptides: antimicrobial, immunomodulatory and anti-inflammatory effects. In Diet, immunity and inflammation. P.C. Calder and P. Yaqoob (ed). Woodland Pub. Limited, 80 High Street, Sawston, Cambridge UK
Smith, N.S., M.L. Oelschlager, M.S.A. Rasheed, R.N. Dilger. 2020. Dietary soy isoflavones reduce pathogen-related mortality in growing pigs under porcine reproductive and respiratory syndrome viral challenge. Journal of Animal Science, 98(4), February 2020. https://doi.org/10.1093/jas/skaa024.
Cemin, H. S., H. E. Williams, M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. Derouchey, K. F. Coble, B. A. Carrender, and M. J. Gerhart. 2020. Estimate of the energy value of soybean meal relative to corn based on growth performance of nursery pigs. J. Anim. Sci. Biotech 11:70. https://doi.org/10.1186/s40104-020-00474-x
Demas, G., T. Greives, E. Chester and S. French. 2011. The energetics of immunity. In Ecoimmunology. G. Demas and R. Nelson. Chap. 8. Oxford Univ. Press. Univ. Oxford, England.
Lewis, A.J. 2001. Amino acids in swine nutrition. In Swine Nutrition, 2nd edition. A.J. Lewis and L.L Southern (ed). CRC Press LLC, 2000 N.W. Corporate Blvd, Boca Raton Florida USA 33431.
Boyd, R.D. and D.E. Bauman. 1989. Mechanisms of action for somatotropin in growth. In: D.R. Campion, G.J. Hausman & R.J. Martin (Eds.) Animal Growth Regulation. Chapter 12, pp. 257-293. Plenum Press, NY.
Decuypere, E. and J. Buyse. 2005. Endocrine control of postnatal growth in poultry. J. Poul. Sci. 42:1-13.
National Research Council (NRC). 1987. Predicting feed intake of food-producing animals. National Academy Press, Washington, DC, USA.
Tillman, P.B. 2019. Determination of nutrient values for commercial amino acids. J. Appl. Poult. Res. 28:526-530. https://doi.org/10.3382/japr/pfz010
R. Dean Boyd, PhD, is an adjunct professor of Animal Nutrition at North Carolina State U. and Iowa State University.; Mike Johnston, MS, is an animal nutritionist and research scientist, United Animal Health; James L. Usry, PhD, is research scientist and principle managing partner of Ani-Tek Group LLC; Richard E. Austic, PhD, is emeritus professor of poultry nutrition at Cornell University.
Acknowledgments
This paper is based on research conducted by the Pig Improvement Co. USA (Technical Memo 211 by Johnston et al. 1999) and published as a research abstract (JAS 77 Suppl. 1:69, 1999). We acknowledge the informal review of diets and responses by Drs. David H. Baker (deceased) and Norlin Benevenga following the ASAS presentation in 1999. This work was presented in part at the John F. Patience International symposium on swine nutrition, August 2021.Download printable PDF.