Classifying maternal resilience for improved sow welfare, offspring performance
Small differences at an early age may translate to impactful differences in the mature market hog.
By Andrea M. Luttman and Catherine W. Ernst
Group-housing sows during gestation is not a new concept from a global perspective. A search for academic articles on the topic of gestational group-housing reveals thousands of results with many articles specifically looking at social stress in gestating sows and prenatal stress of the offspring. We have a lot of information on how gestational group-housing affects the sows as well as their offspring, but the swine industry also faces a great challenge in that it is not possible to completely remove social stressors.
Not only can we not fully remove social stressors, but it is also difficult to optimally manage social stress in gestating sows. This is due to many factors including the sow’s complex social nature as well as limitations in management arising from facilities, infrastructure, labor and overall cost. An individual animal’s response to group-housing depends on their social cohort and their ranking within the social hierarchy of that specific cohort.
In a facility that operates with batch farrowing, groups of gestating sows may be stable, whereas those that operate with continuous farrowing might deal with dynamic groups. Dynamic groups, where animals are added or removed periodically, results in disruption of the social hierarchy and leads to social stress while the hierarchy is reestablished.
Additionally, the type of group-housing facility will impact pressures of social stress through competition for resources and facilitation of evading aggression. Use of electronic sow feeders (ESF) allows individual management and non-competitive eating environment but may still cause perceived competition for resources resulting in queueing at the feeder gate and increased aggression.
Floor pen feeding requires less infrastructure and may be more cost-effective to convert to but creates a competitive eating environment. Large, open pens do not allow sows to escape an aggressor. Adding partition walls may help reduce duration of aggressive interactions by providing a physical barrier for a sow to escape behind but increases cost. Thus, identifying strategies for mitigating effects of social stress in gestating sows is critical.
Social stress not only negatively impacts the sow’s welfare, but it impacts her production ability and her offspring. It’s been reported that gestational stress results in a higher number born alive, decreased birthweights, increased pre-weaning mortality, altered stress response, reduced sickness behavior and abnormal maternal behavior.1-7 Despite our knowledge of the problem, we have yet to find a sustainable solution. It is for this reason that our lab began researching maternal resilience to social stress. When we cannot fit the environment to each individual animal to minimize social stress, we must select for animals that best fit the environment. By selecting for resilience, the ability to be minimally affected by or quickly recover from stressors, we could improve sow welfare and reduce prenatal exposure to social stress.
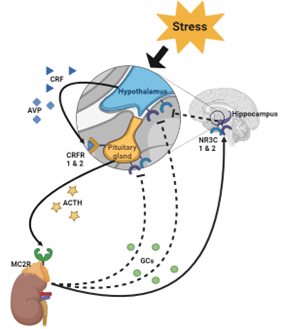
In our research working with gestating sows, we focus on ability to quickly recover from stress by observing the natural cortisol response patterns following social mixing. We have found a high degree of individual variation of baseline levels and degree of acute response. This variation likely originates from underlying molecular differences of the hypothalamic-pituitary-adrenal (HPA) axis. The HPA axis is a complex system that involves the hypothalamus, pituitary gland, adrenal gland, hippocampus, and amygdala. As summarized in Figure 1, the HPA axis primarily relies on the well understood hormone cascade of the hypothalamus secreting corticotropin releasing factor (CRF) which is bound to its receptor on the pituitary gland to stimulate secretion of adrenocorticotropic hormone (ACTH) which travels to and binds its receptor on the adrenal gland to stimulate secretion of the glucocorticoid (GC) cortisol.
However, it is often overlooked that the HPA axis is also a negative feedback loop—cortisol has receptors in the hippocampus, hypothalamus, and pituitary gland which have an inhibitory effect when bound to cortisol. By focusing on the animal’s ability to recover from stress (returning to baseline levels) rather than degree of HPA activation, we are estimating the capacity of the animal’s HPA axis to self-regulate. Conversely, we assume that failure of the HPA axis to activate or return to baseline levels is a result of HPA dysregulation.
In our recent study we measured salivary cortisol from 64 first and second parity sows at an acute social mixing event to identify patterns of stress resilience (SR) or stress vulnerability (SV). In short, sows were bred and confirmed pregnant by ultrasound at approximately 22-24 days of gestation (dg). The pregnant sows were moved from individual breeding stalls to a free-access stall system (FAS) at 25dg.
Sows were placed into groups of 8 sows/pen (8 pens, n=64) and kept contained in individual stalls within the FAS until released for an acute mixing at 30dg. Saliva was collected at -1, 0, +1, +2, +3, and +7 days relative to the acute mixing (i.e. 29, 30, 31, 32, 33, 37dg). Samples were collected at noon of each day by suspending a clean cotton rope at the front of each individual stall and allowing the sow to chew on the rope until just saturated . Ropes were then centrifuged in Salimetrics collection tubes (Salimetrics LLC, Carlsbad, CA) to extract the saliva. Salivary cortisol was measured using a commercially available, high-sensitivity ELISA kit.
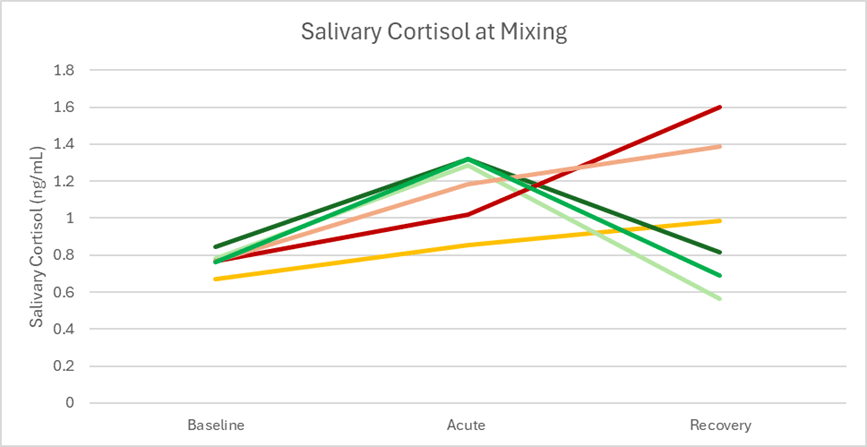
Salivary cortisol levels were used to examine each sow’s response pattern over time, categorized into a baseline stage, acute stage, and recovery stage. Focal sows were identified as having a stress resilient or stress vulnerable pattern, modeled after our earlier study examining resilience to weaning stress (Luttman et al, 2023). Resilient sows exhibit a strong acute response with complete recovery to baseline by 7d post-mixing; vulnerable sows exhibit moderate acute response but cortisol continually elevates suggesting a chronic-stress pattern (Figure 2).
Of the 64 sows, we identified 9 SR and 9 SV sows to follow to farrowing. Litters had 15 +/- 3 total born (range, 9-20), with 12 +/- 3 born alive (range 7-18). We observed no statistical differences in total born or number born alive between SR and SV litters. Within 24 hours of birth, 1 female and 1 male from each litter was euthanized for terminal sampling. One SV litter had to be removed from study due to the sow laying on piglets, falling below our study minimum requirement 2 males and 2 females per litter. All remaining piglets (n=158, 9 +/- 3 per litter, range 4-14 per litter) were used to characterize offspring differences as measured by birthweight, weaning weight, and suckling stage average daily gain (ADG). Birth weight was recorded at 1d post-farrowing during processing. Individual weaning weights were recorded at 1d pre-weaning (26 +/- 4d of age). Suckling ADG was calculated by subtracting the birth weight from the wean weight and dividing by the weaning age.
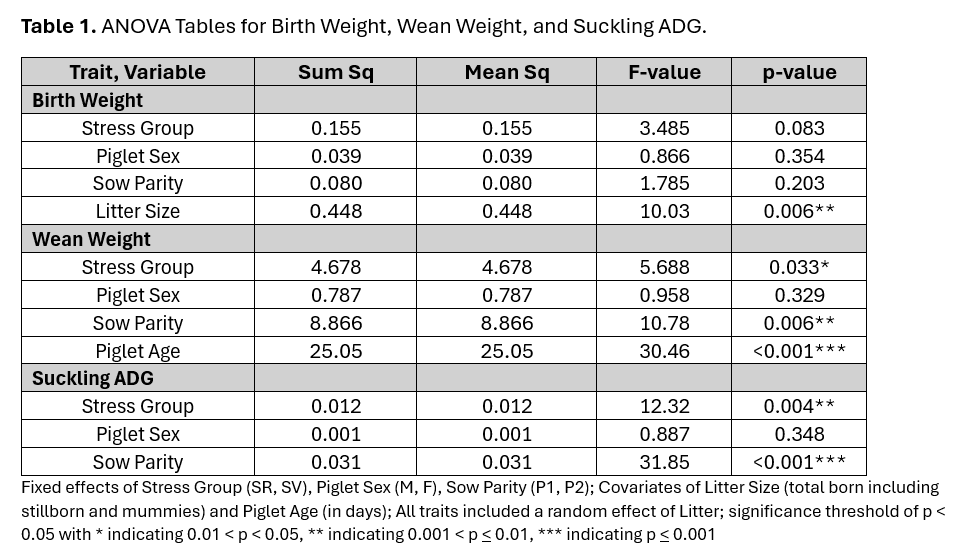
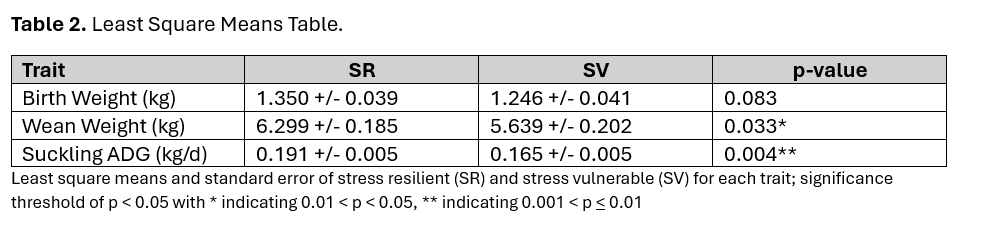
Analysis of each trait was performed using Gaussian linear mixed models. Each analysis included the random effect of litter and fixed effects of stress group (SR or SV designation of the sow), sow parity (P1 or P2), and sex of the piglet (M or F). Birth weight was corrected for the total number of piglets produced by the sow using a covariate of Litter Size. Wean weight was corrected for age at weaning using a covariate of Piglet Age. Effect of each variable is summarized in Table 1. The impact of litter size on birth weight is to be expected, with increasing litter size being associated with lower birth weights. Sows in their second parity had piglets exhibiting a higher daily gain and wean weight. This is also to be expected as first parity sows are still growing and perhaps unable to provide as much nutrition to their piglets. Piglet sex was not significant for any of the three traits. Regarding maternal resilience, we observed a tendency for SR-born piglets to weigh more at birth than SV-born piglets, and significant differences in daily gain and weaning weight (Table 2).
Sows that exhibited resilience to gestational social stress tended to give birth to piglets that were 0.10 kg heavier on average. These SR-born piglets then gained 0.03 kg more per day leading to a weaning weight that was 0.67 kg greater on average when compared to SV-born piglets. While these differences may not seem meaningful at a glance, there is extensive research showing that birthweight and pre-weaning performance is a crucial indicator of survivability and later growth performance. Small differences at this early age may translate to impactful differences in the mature market hog. It is for this reason that we are continuing to follow these offspring to observe their post-weaning performance.
The post-weaning performance of these pigs will also help us discern if the pre-weaning differences between SR- and SV-born piglets are due to sow-factors or piglet-factors. If differences in growth performance disappear following weaning, it guides our understanding of how gestational stress is impacting the offspring. Perhaps stress in early-mid gestation could be influencing the lactation ability of the sows leading to the differences in piglet growth. If differences in growth persist post-weaning, it suggests underlying differences in the piglets such as fetal programming that occurred as a result of exposure to prenatal stress. Regardless of the exact mechanism through which maternal resilience is altering piglet performance, our lab is very excited about these results and looks forward to continuing the characterization of stress resilience in pigs and elucidating the underlying molecular mechanisms that shape the resilient pig.
Author information and acknowledgementsAML is a PhD candidate in Genetics & Genome Science and CWE is a Professor of Animal Science at Michigan State University. We would like to thank the MSU Swine Teaching and Research Center for their continued support of our research.
This project is funded by USDA Agriculture and Food Research Initiative (award number 2022-67015-36865) and AML is supported by a pre-doctoral fellowship (award number 2022-67011-36562) from the USDA National Institute of Food and Agriculture.
References1. Couret, D., Jamin, A., Kuntz-Simon, G., Prunier, A., & Merlot, E. (2009). Maternal stress during late gestation has moderate but long-lasting effects on the immune system of the piglets. Veterinary Immunology and Immunopathology, 131(1–2), 17–24. https://doi.org/10.1016/j.vetimm.2009.03.003
2. Couret, D., Prunier, A., Mounier, A. M., Thomas, F., Oswald, I. P., & Merlot, E. (2009). Comparative effects of a prenatal stress occurring during early or late gestation on pig immune response. Physiology and Behavior, 98(4), 498–504. https://doi.org/10.1016/j.physbeh.2009.08.003
3. Jarvis, S., Moinard, C., Robson, S. K., Baxter, E., Ormandy, E., Douglas, A. J., Seckl, J. R., Russell, J. A., & Lawrence, A. B. (2006). Programming the offspring of the pig by prenatal social stress: Neuroendocrine activity and behaviour. Hormones and Behavior, 49(1), 68–80. https://doi.org/10.1016/j.yhbeh.2005.05.004
4. Kranendonk, G., Hopster, H., Fillerup, M., Ekkel, E. D., Mulder, E. J. H., & Taverne, M. A. M. (2006). Cortisol administration to pregnant sows affects novelty-induced locomotion, aggressive behaviour, and blunts gender differences in their offspring. Hormones and Behavior, 49(5), 663–672. https://doi.org/10.1016/j.yhbeh.2005.12.008
5. Kranendonk, G., Hopster, H., Fillerup, M., Ekkel, E. D., Mulder, E. J. H., Wiegant, V. M., & Taverne, M. A. M. (2006). Lower birth weight and attenuated adrenocortical response to ACTH in offspring from sows that orally received cortisol during gestation. Domestic Animal Endocrinology, 30(3), 218–238. https://doi.org/10.1016/j.domaniend.2005.07.001
6. Lay, D. C., Kattesh, H. G., Cunnick, J. E., Daniels, M. J., McMunn, K. A., Toscano, M. J., & Roberts, M. P. (2008). Prenatal stress effects on pig development and response to weaning. Journal of Animal Science, 86(6), 1316–1324. https://doi.org/10.2527/jas.2007-0812
7. Lay, D. C., Kattesh, H. G., Cunnick, J. E., Daniels, M. J., Kranendonk, G., McMunn, K. A., Toscano, M. J., & Roberts, M. P. (2011). Effect of prenatal stress on subsequent response to mixing stress and a lipopolysaccharide challenge in pigs. Journal of Animal Science, 89(6), 1787–1794. https://doi.org/10.2527/jas.2010-3612
Luttman is a PhD Candidate & USDA NIFA Pre-Doctoral Fellow in the Genetics & Genome Sciences program and Ernst is the chair of the Department of Animal Science, both at Michigan State University.