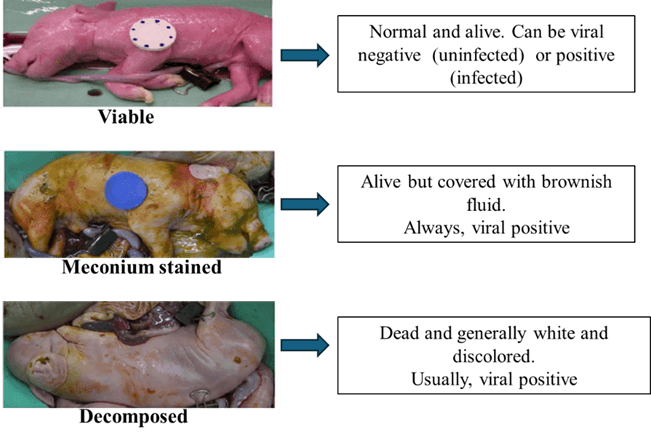
Investigations into fetal response to PRRSV infection
Outcomes of pigs born from PRRSV infected mothers are variable in terms of infectivity, survivability and thriftiness.
By Kristen E. Walker and Joan K Lunney
Porcine reproductive respiratory syndrome (PRRS) is the costliest disease for U.S. pig farmers, with estimates of $664 million in losses to the US pork industry considering both breeding and growing herds. In PRRS virus infected gilts late gestation abortions are observed, or if the gilts succeed in birthing, they deliver weak, stillborn or mummified piglets as well as apparently healthy piglets. These effects are indiscriminate and the reasoning for why some fetuses are more severely impacted by the virus is not fully understood.
Outcomes of pigs born from PRRSV infected mothers are variable in terms of infectivity, survivability and thriftiness. They can range from viable piglets that are uninfected, or infected with low or high amounts of virus, or viable piglets that are infected with virus and stained with a brownish-yellowish fluid called meconium (MEC), or non-viable/dead piglets (Figure 1).
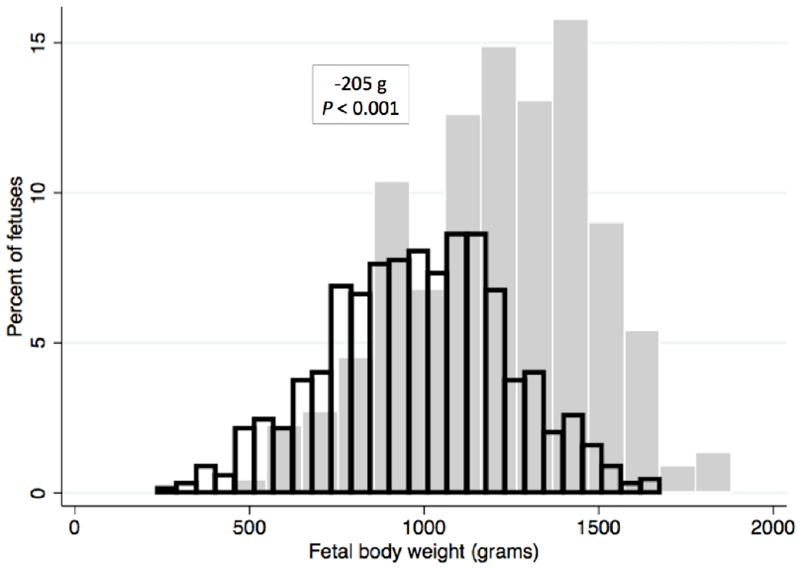
MEC stained fetuses are expected to die within a short time if born alive. Fetal body weight is affected by late gestation infection, with viable fetuses from PRRSV-infected gilts weighing less than those from control gilts (Figure 2). Many viable fetuses from PRRSV infected gilts are less thrifty than their counterparts born from non-infected gilts.
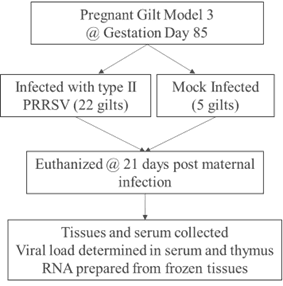
To detail the phenotypic effects on the fetus and provide information about the genomic basis for differences in fetal responses to congenital PRRSV infection, the pregnant gilt infection model (PGM) was developed by the Harding lab (Figure 3). Gilts from a PRRSV free high health herd, were synchronized to estrus, followed by fixed time artificial insemination. The PGM was carried out using the guidelines of the Canadian Council of Animal Care and with approval of the University of Saskatchewan’s Animal Research Ethics Board (Protocol #20180071). Pregnant gilts were transferred to the University of Saskatchewan level 2 biocontainment facility late in gestation. On gestation day 85, 22 gilts were inoculated PRRSV isolate NVSL 97–7895, while five gilts were mock infected with medium. At 21 days post maternal infection (dpmi) pregnant gilts and their fetuses were euthanized, and tissues and serum collected and frozen for future analyses.
Using information gathered from the PGM studies, our research group has designed experiments to explore gene expression data with the goal of identifying PRRSV resistant pigs which then could lead to the development of PRRSV resistant swine herds. To provide information on which genes are turned on or off in these varied fetal conditions following PRRSV infection, our group is interested in assessing tissues from viable fetuses and evaluating tissue gene expression. Gene expression studies provide crucial insights into how cells and tissues function by revealing which genes are being activated or deactivated following PRRSV infection. This will assist in understanding the molecular mechanisms behind PRRSV resistance/susceptibility, virus transmission and disease progression.
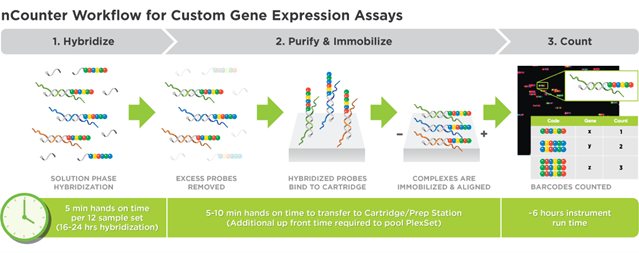
Methods: Experiment 1To explore the relationship between gene expression, susceptibility and resistance using the PGM, we used next generation NanoString Technology. This technology uses reporter and capture probes to detect unique barcodes for each target gene; output is individual gene counts (Figure 4). NanoString’s targeted transcriptomics evaluates RNAs and is more quantitative and sensitive, with improved signal to noise and increased statistical power, than other methods such as RNAseq and qPCR. For our analyses we created custom pig gene targeted codesets to investigate gene expression and thus pathways of biological pathways and functions. This sheds light on which genes and potential traits to selectively target for identifying potential PRRSV resistant breeding stock. All of our studies were based on analyses of RNA that was carefully extracted from frozen PGM3 tissues.
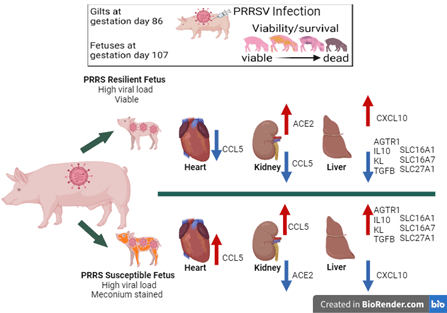
In our first round of gene expression studies, we were interested in two categories of viable fetuses: those with high viral load (HVL_VIA), resilient fetuses, and those with high viral load and meconium staining (HVL-MEC), susceptible fetuses. The study was intended to investigate which genes are altered when a PRRSV infected fetus contracts the virus, acquires a high viral load and progresses to MEC staining.
For our studies gene expression and biological pathways were targeted based on their hypothesized functions during PRRSV infection in three critical tissues: heart, kidney and liver. The heart's main function is to pump blood throughout the body; kidneys are essential for removing waste products, maintaining fluid and electrolyte balance, and regulating vital bodily processes; and the liver filters all of the blood in the body and breaks down and carry away waste. We selected these vital organs because we had particular interest in vascular regulation, associated with the renin-aldosterone-angiotensin system (RAAS) following infection.
The RAAS is key in maintaining homeostasis through regulation of blood vessel dilation and constriction, ion and water transport mechanisms systemically, and affects the development of new blood vessels, cell proliferation, growth, differentiation and apoptosis, tissue inflammation, and hormonal secretion affecting systems in the heart, kidney and liver.
We hypothesized that there was a difference in gene expression affecting the biological pathways of interest in the heart, kidney, and liver of those fetuses that are resilient against the virus (HVL_VIA), and those which are susceptible (HVL_MEC) and assumed would succumb to the PRRSV if the infected gilt was allowed to reach full term gestation.
Results: Experiment 1Results showed that several genes were impacted at different levels in the fetal tissues following PRRSV infection, which may influence the fetus’ ability to resist or succumb to infection (Figure 5). In the heart, and kidney chemokine CCL5 was decreased in the resilient fetus but increased in the susceptible fetus. CCL5 is an immune gene which functions to attract immune system cells to sites of inflammation and has been shown to play a role in blood vessel formation. In the kidney ACE2 is increased in the resilient and decreased in the susceptible fetuses; ACE2 which affects new blood vessel formation and blood pressure regulation in the PRRSV infected fetus.
The liver of the PRRSV infected fetuses showed the most gene expression changes with varied effects in immune genes, RAAS genes and solute carriers (Figure 5). Solute carrier genes (SLCs) transport various molecules: ions, nutrients, and metabolites across cell membranes, moving them from one side of the membrane to the other. These transporters play a critical role in maintaining cellular homeostasis and physiological functions like nutrient uptake. It is possible that during fetal PRRSV infection there is an impact on the fatty acid and glucose transport affecting energy production and usage resulting positively resilience or negatively in fetal demise.
Methods: Experiment 2Among the variability in the fetal response to maternal PRRSV infection there remain some resistant fetuses which do not contract the virus at all, or when contracted, the infection levels remain minimal, below 5 log of virus (low viral load, LVL). Our second study was designed to investigate the biological pathways activated when the fetus of a gilt has detectible levels of virus in the placenta, and serum but resists progressive viral infection in the thymus (above 4.99 log virus) or meconium staining. We were interested in the fetal thymus because it is known to be a site of replication for the virus. There was interest in the heart as a follow up to the results of our previous study, and because the heart is arguably the most vital organ.
We hypothesized that there are many functions at play to protect the resistant (LVL) fetus including activation of the innate immune system, effects on transport of molecules across the cellular membrane, energy metabolism, and tissue-specific stress related effects in the thymus and the heart. Considering the COVID-19 pandemic there we also incorporated target genes associated with SARS-Cov2 infection results as this virus and PRRS virus are members of the same viral order – Nidovirales. Both PRRSV and SARs-CoV2 are enveloped, positive-stranded RNA viruses which infect vertebrates. We also expanded the category of tissue specific stress response genes from our first study and included cell surface markers associated with PRRSV infection in the Viral Reactive category.
Results: Experiment 2Gene expression results indicated little to no differential gene expression in the thymus. Only 4 of 179 target genes were differentially expressed (not shown). Studies have shown that the thymus is one of the major sites of PRRSV replication and is known to be an immune tissue important for lymphatic function. In our study however, there was no differential expression until the virus reached the thymus (HRT_THYL), but even then, only a minimal gene expression response was observed.
This may be due to thymic attempts at protecting itself from a cytokine storm, which is a severe immune response that happens when the body releases too many cytokines into the blood at once. Cytokines storms have been observed in the human body’s reaction to SARS-Cov2 resulting in severe complications including death. Since the LVL fetuses in this study remained resistant to PRRSV we suppose that the suppressed gene expression in the thymus could provide protection to a cytokine overreaction that would be devastating to the fetus.
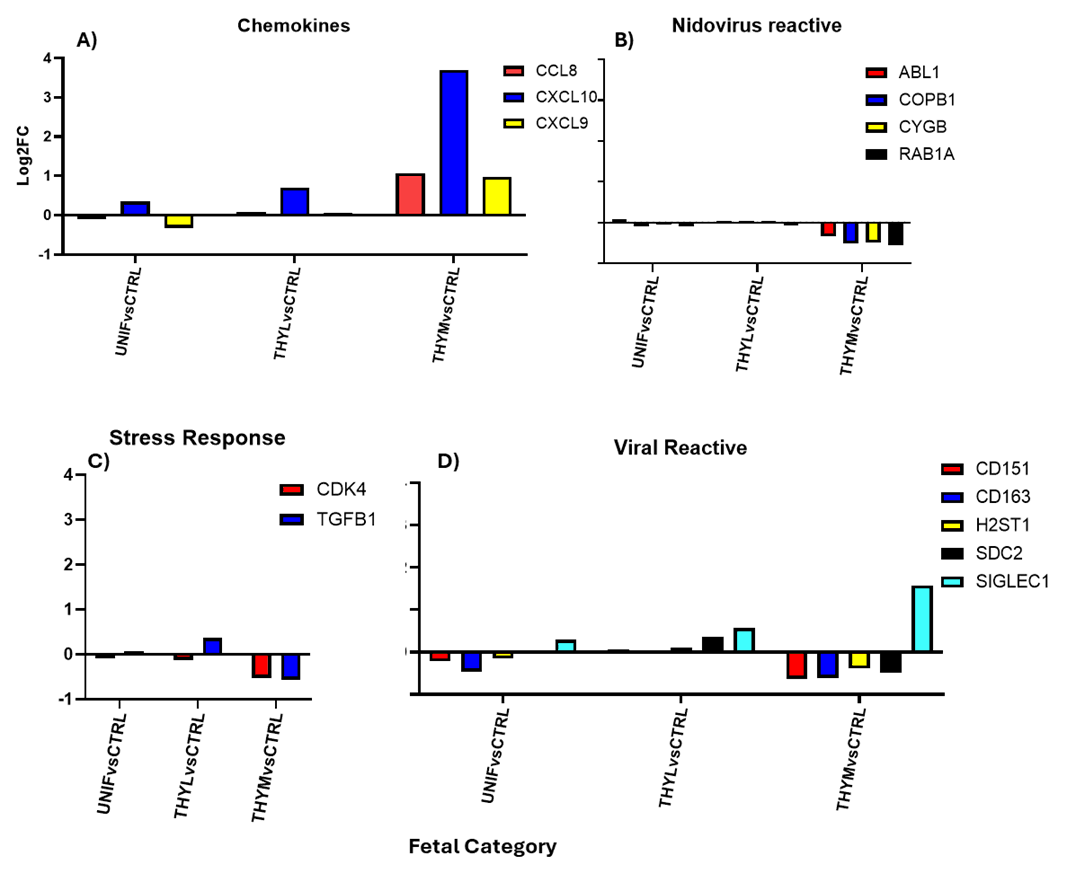
NanoString codeset gene expression levels were calculated for each gene in each category. Graphs show comparative fold changes as log2 fold change (log2FC) for UNIFvsCTRL, THYLvsCTRL, THYMvsCTRL as compared to CTRL RNA. All results shown are of significantly differential expression as adjusted *P.<0.05. A) Chemokine genes; B) Nidovirus reactive genes; C) stress response; D) viral reactive genes.
In the heart, robust expression in over 30 of 179 genes was observed, but only after the virus reached detectable levels in the infected fetal thymus (HRT_THYL) (Figure 6). No differential gene expression was detected when the virus was only in the placenta and serum, only when the virus reached the fetal thymus were there significant gene expression differences. An increase in several immune gene categories was observed. The chemokines (Figure 6A) in LVL fetuses which support the idea that these immune cells are mobilized to tissues which are experiencing stress in the presence of virus.
There was a general decrease in SARS-CoV2 associated genes in the HRT (Figure 6B) which may give more information about the mechanisms of the fetuses’ ability to resist progression to greater levels of pathogenicity. We noted a decrease in stress response genes (Figure 6C) in the HRT once the virus reached the thymus; this was observed in our previous study.
For Virus Reactive genes, we observed an increase of SIGLEC-1 (Figure 6D) which is known to be involved in the receptor-mediated endocytosis entry process of PRRSV into porcine alveolar macrophages and is associated with virus attachment and internalization. There was decreased gene expression in the heart of fetuses when the virus reached the fetal thymus which confirms SIGLEC-1 as a r4levant PRRSV receptor molecule. However, these fetuses did not progress to more detrimental viral levels which may correlate to the decrease of the obligate PRRSV receptor, CD163 (Figure 6D).
This may correlate with fetal PRRSV resistance in those fetuses. It is possible that the differential expression of these cell surface markers CD163, CD151, and CD169 following PRRSV infection in this study may indicate tissue specific shifts in susceptibility and numbers of macrophage populations (Figure 6D). We think that this may be indicating resistance to progressive infection by activating innate immune networks and employing mechanisms to recruit important defensive cells, redirect energy and decrease expression of possible PRRSV receptors to protect its most vital organ.
ConclusionIn conclusion, our gene expression results provide clues into the mechanism of fetal susceptibility and resilience to congenital PRRSV infection in fetuses. Genes related to immunity, angiogenesis, solute transport and stress response are modulated in response to fetal PRRSV infection. They either protect the fetus from progressing to, or provide futile attempts at rescuing the fetus from, advanced levels of infection.
Future work will focus on expanding the knowledge of which genes, and their mechanistic pathways, are most affected by in-utero infection. These will provide important targets for efforts to influence fetal survival outcomes, to breed for more thrifty and virus resilient pigs.
Walker is a laboratory manager/support scientist and Lunney is a distinguished senior research scientist, both with USDA's Agricultural Research Service.