Determining probiotic mechanisms, investigating MGB in weaning pigs
By Sheila Jacobi
Between the gut and the brain, there are complex bidirectional communication pathways called the microbiota-gut-brain (MGB) axis. The gut and the brain are constantly communicating and influencing each other through neural, hormonal and immune signals. The communication is integral to physiological homeostasis within an animal.
The gut microbiota has been linked to the homeostasis of gut health and efficiency of digestion and absorption of nutrients. More recently the gut microbiota and microbial endocrinology have become a major focus of research in animal health and disease because of the many links to the animal well-being. Whereby, a diverse and stable gut microbiota has been correlated with improvements in immunological, physiological and neurological states across life span.
The composition of the gut microbiota is closely associated with nutrition, environment and microbial genomics. Microbial derived neurochemicals such as serotonin, gamma-aminobutyric acid (GABA), dopamine, norepinephrine and acetylcholine are implicated in regulation of appetite, metabolism and behavior. In fact, the gut and its microbiota are a major source of neurochemicals which links it to neural function. Additionally, gut hormones that regulate appetite control in the brain have been shown to be modulated by microbiota-derived hormones that regulate feed intake in other animals.
The food animal production industry has used probiotics to improve gut health as an alternative to in-feed antibiotics to help maintain gut health and performance. While many defined probiotics are available for use, we still have inconsistent results of probiotics depending on the health status or the environment in which they are used. Therefore, defining probiotic mechanisms could be advantageous in modulation of the MGB axis to improve animal performance and well-being.
MGB axis importanceProbiotics are defined as a living microorganism that when supplemented in adequate amounts confers a health benefit to the host (Butel, 2014). Through interacting with the host microbiota and intestinal epithelium, probiotics have been shown to exert a wide range of effects upon host health, with various strains improving metabolism, immunity, endocrine function and intestinal health (Salami, 2021). Perhaps the most intriguing effects of probiotics on the host is their modulation of brain physiology and behavior. Considerable research over the last decade has documented how probiotics can influence various central nervous system processes which affect appetite, mood and behavior (Yong et al., 2020).
In human research, the term psychobiotics are now defined as microbiota-targeted interventions such as probiotics that influence bacteria-brain relationships or the MGB axis (Yong et al., 2020). As the evidence to support the effects of psychobiotics on brain and behavior continues to develop (Yong et al., 2020), the field is now turning to mechanistic studies to elucidate the biological underpinnings of the MGB axis.
In livestock, work by Cheng et al. (2019) in poultry investigated the effects of Bacillus subtilis supplementation in aggressive hens and was able to show a reduction in feather pecking and kicking through intestinal modulation of serotonin synthesis and signaling. Further, others have shown microbial neurochemicals associated with probiotic supplementation regulate gut hormones that modulate brain feed intake centers in weaning pigs (Sun et al., 2021).
Therefore, a novel focus in enteric microbial research has shifted towards neurophysiology and immune modulatory effects of the gut microbiome, and much of this interaction revolves around the MGB axis. The intersection of microbiology and neurophysiology, known as microbial endocrinology, holds that disruptions to the gut microbiome and intestinal health can influence neural physiology, cognition, behavior, mood and feed intake (Rogers et al., 2016). Microbial endocrinology-based mechanisms mediate the ability of bacteria and host to interact with each other in a bidirectional manner, ultimately influencing host physiology ranging from susceptibility to infectious disease and behavior to overall host performance (Lyte and Lyte, 2019).
We know dietary probiotics alter composition and modulate global metabolic function of the intestinal microbiome (Plaza-Diaz et al., 2019). What we do not completely understand are the bio/neurochemical mechanisms through which probiotics modulate the MGB axis. The psychophysiological response to stress, which affects many aspects of an animal’s physiology and ultimately performance of the animal, should be of primary interest to the food animal industry because in addition to performance it is linked to overall animal well-being. The objective of this work is to understand the mechanisms by which two genome-sequenced probiotics improve intestinal health, and therefore could impact overall weaning pig performance. The first phase of this work was to determine probiotic mechanisms in vitro on intestinal barrier function using pig intestinal cells and phase two is to investigate the MGB in weaning pigs.
Biology of probiotic strainsTwo novel probiotic strains Pediococcus acidilactici-3A and Lactobacillus helveticus-4A were isolated and genome-sequenced at the Molecular and Cellular Imaging Center in Wooster, Ohio in collaboration with Rafael Jimenez-Flores of the Department of Food Science and Technology at The Ohio State University. In our research group, we have isolated a particular P. acidilactici-3A strain from Gouda cheese. This strain stood out over 137 other LABs, in terms of lipolytic, proteolytic and neurochemical synthesis (García-Cano et al., 2019; in preparation).
In terms of genetic analysis, the strain has a genome sequence of 1,947,223 bp. According to gene prediction and functional annotation the probiotic has 1,956 genes. Gene annotation elucidated possible neurotransmitters production such as GABA by L-Glutamate via glutamate decarboxylase activity. As well, serotonin production via L-Tryptophan metabolism using a tryptophan hydroxylate to produce L-5-hydroxytryptophan (5’HT) and an aromatic L-amino acid decarboxylase to generate serotonin. Also, we have elucidated norepinephrine (NE) production and dopamine (DA) production via Phenylalanine/Tyrosine metabolism, using a tyrosine oxidase and an aromatic L-amino acid decarboxylase (García-Cano et al, in preparation).
Lactobacillus helveticus-4A is a lactic acid bacterium. We have sequenced the complete genome and it showed 1,834,843 bp in its genome with four genes with putative function of b-galactosidase were found that are important in lactose metabolism (García-Cano et al, 2022). We further elucidated the mechanism of production of neurotransmitters 5-HT, NE and DA, through gene sequences associated with the same metabolic pathways described previously for Pediococcus acidilactici.
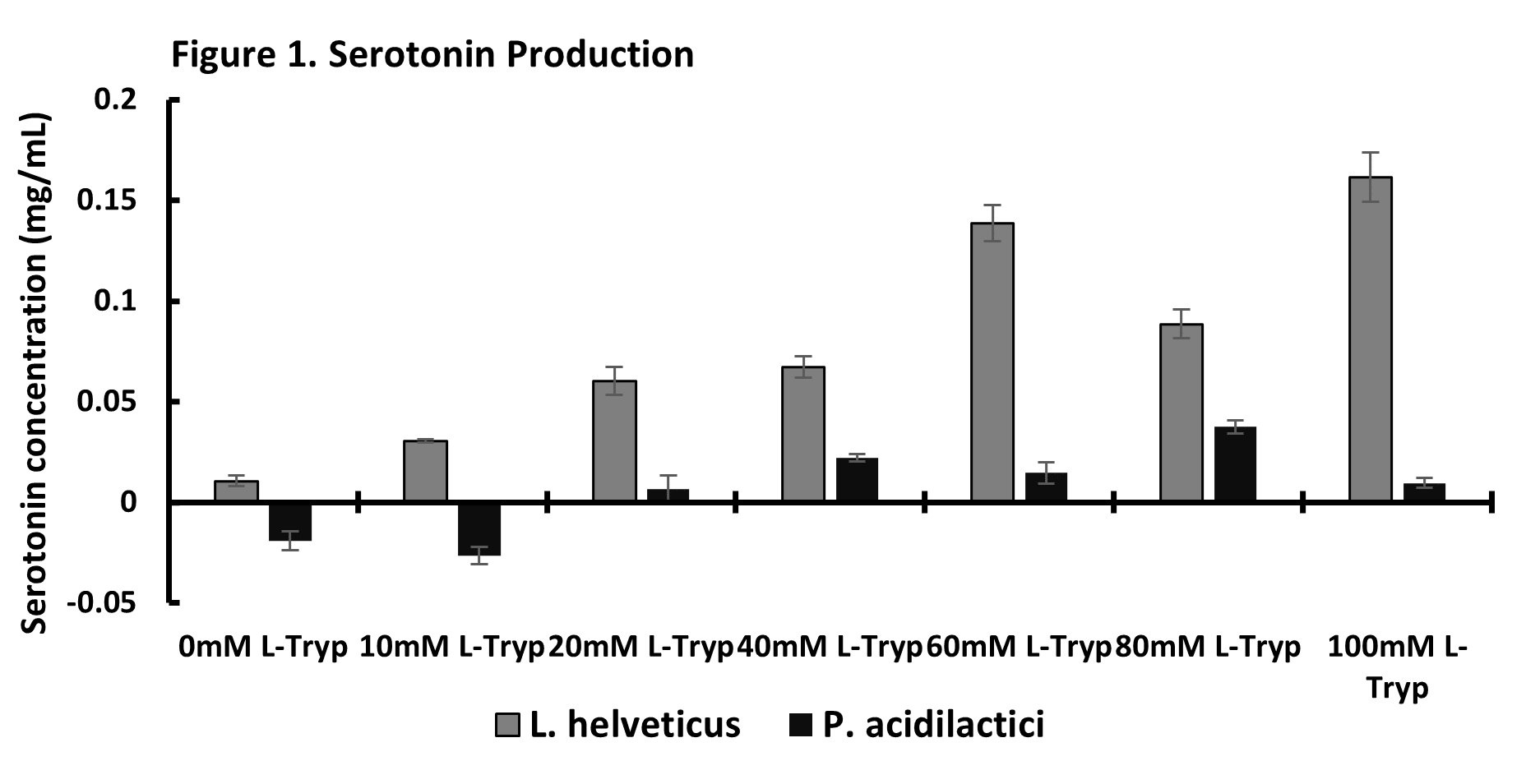
Neurotransmitter secretionGrowth of Lactobacillus helveticus-4A with increasing concentration of L-tryptophan in the culture media showed increased secretion of serotonin by the probiotic (Figure 1). Pediococcus acidilactici-3A did not increase serotonin secretion in the presence of increasing concentration of L-tryptophan. Glutamate supplementation into the probiotic growth media preliminarily suggest Pediococcus acidilactici increases GABA secretion, but Lactobacillus helveticus under the same conditions does not increase GABA secretion (data not shown).
Further testing is being done to determine nutrient exposures which alter neurotransmitter secretions in the probiotics. Together the genome sequencing and probiotic cultures exposed to different nutritional conditions provide information regarding how the probiotic responds to its environment and nutrients which may impact communication between the intestinal microbiota community and pig physiology.
Piglet intestinal epithelial cellsTo understand the probiotic and porcine intestinal cell interactions the use of a neonatal intestinal porcine cell line was used. The IPEC-J2 cells used in this study are porcine intestinal epithelial cells and were originally isolated from a one-day old neonatal piglet intestine (Vergauwen, 2015). Cells were cultured in appropriate conditions to induce cell development. Cells were then exposed to the probiotic bacterial strains and an isolated enterotoxigenic Escherichia coli (ETEC) strain 3030-2: K88ac, LT, STb was kindly provided by Philip Hardwidge of Kansas State University isolated from scouring nursery piglets.
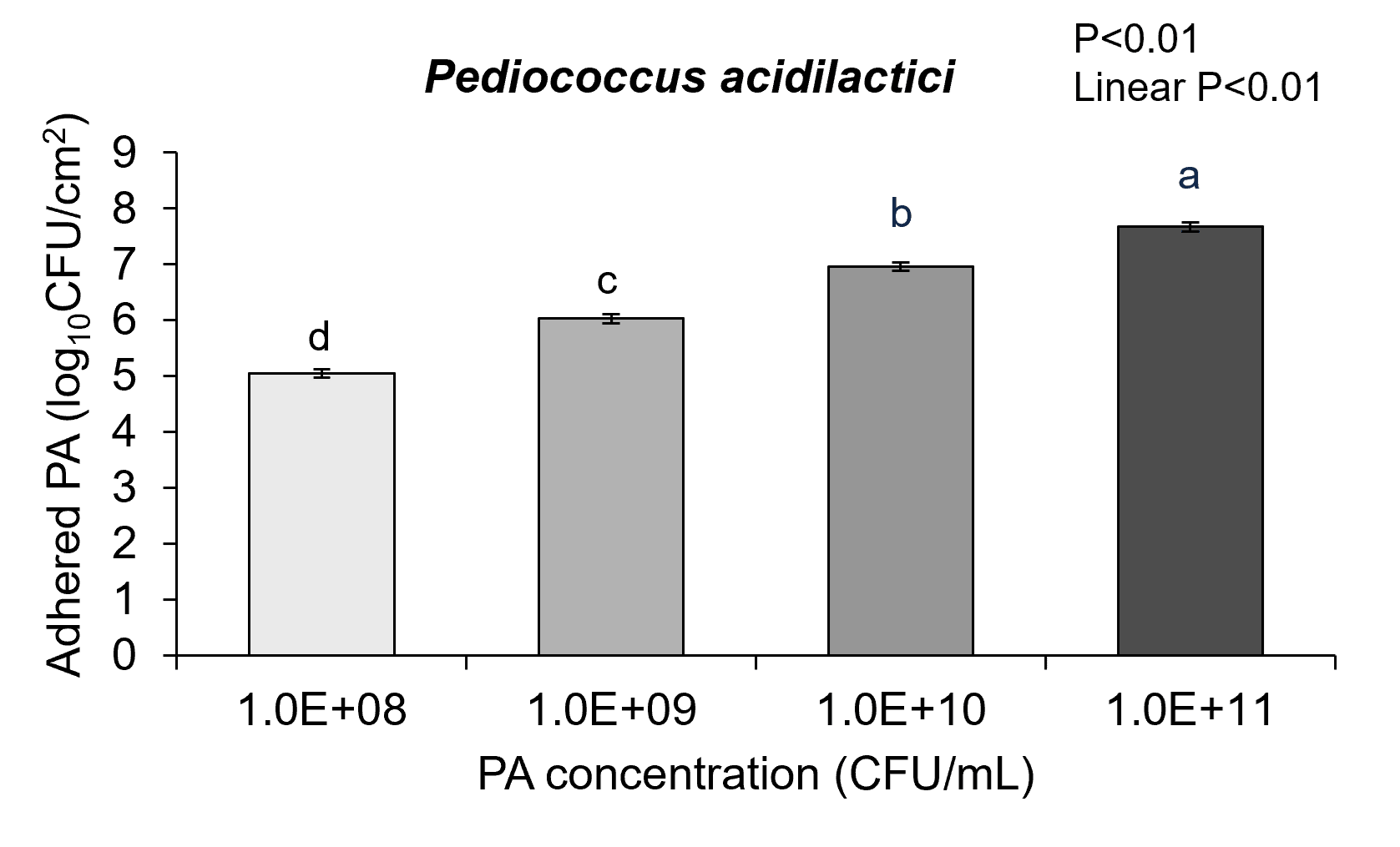
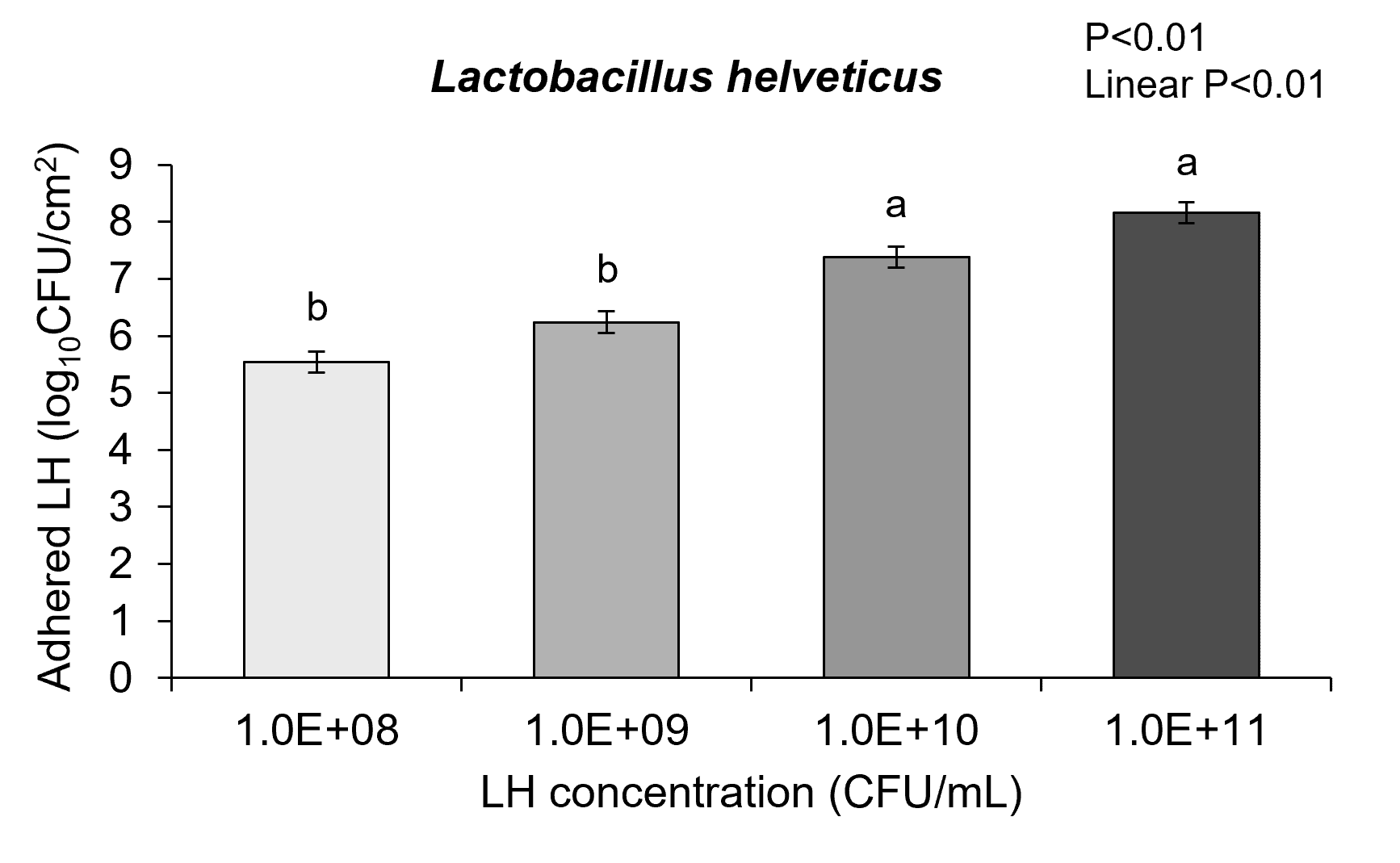
To understand how the piglets’ intestinal cells interact with probiotic bacteria several experiments were performed looking at probiotic adhesion to the intestinal cells and pig intestinal cell health. The IPEC-J2 cells were grown and exposed to each probiotic diluted to appropriate test concentrations. Cells were treated with either Pediococcus acidilactici or Lactobacillus helveticus for 3h at concentrations of 108, 109, 1010, and 1011 colony forming units (CFU)/mL. After treatment with probiotics, plating assays were performed to determine the concentration of probiotics attached to piglet epithelial cells.
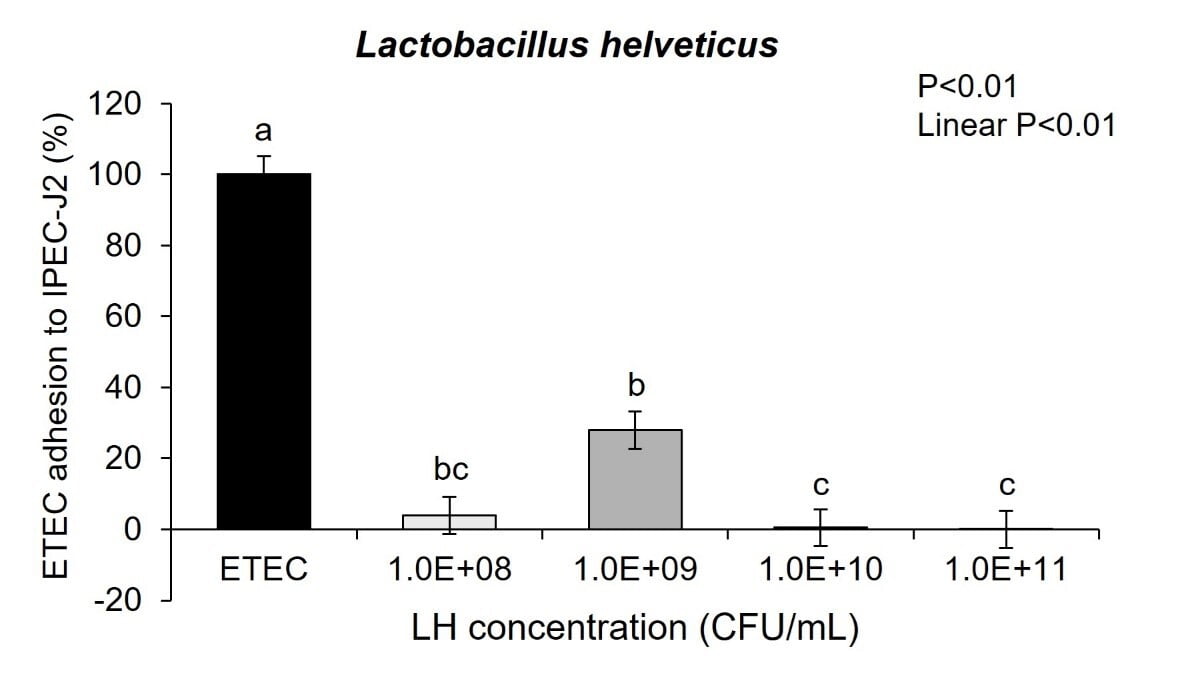
The results of these experiments showed neonatal pig intestinal epithelial cells that were co-cultured with either probiotic had probiotic adhesion to the intestinal cells. Further with increasing concentrations of each probiotic showed there was increased adhesion of Pediococcus acidilactici-3A and Lactobacillus helveticus-4A in a dose-dependent linear response to the intestinal epithelial cells surface (Figure 2a and 2b; linear P<0.01). These adhesion results could significantly impact bacterial-host communication pathways that affect gut health.
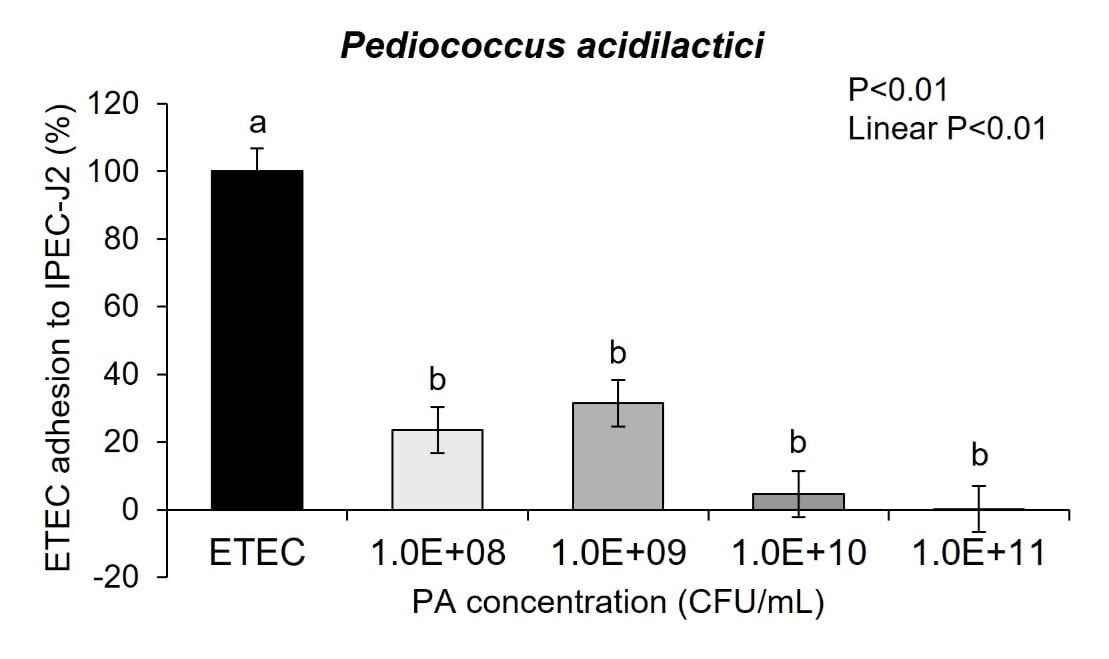
How do probiotics protect cells?To determine if these probiotics could inhibit negative pathogenic effects on the piglet intestinal cells a series of experiments were performed looking at the probiotic and pathogen interactions on the intestinal barrier. Intestinal porcine epithelial cells were cultured under the appropriate conditions and then were treated with either Pediococcus acidilactici or Lactobacillus helveticus at increasing concentrations of 108, 109, 1010 and 1011 CFU/mL and challenged with ETEC. ETEC solution was added at a multiplicity of infection of 2:1 for 3h. Bacterial adhesion was counted according to the procedures of Qiao et al. (2020) with modifications. Samples were serially diluted and plated on Luria-Bertani agar dishes overnight to determine ETEC adhesion. Further, porcine intestinal epithelial cell toxicity was determined following exposure to the probiotics alone or the probiotics + pathogen exposure.
Cell toxicity was measured using a lactate dehydrogenase (LDH) assay (Cayman Chemical, Ann arbor, MI) according to the manufacturer’s protocol. Pediococcus acidilactici-3A and Lactobacillus helveticus-4A were cultured with IPEC-J2 pig intestinal cells in the presence of ETEC for 3 hours. Both probiotics linearly decreased adhesion of ETEC to the IPEC-J2 cells across the probiotic doses of 108, 109, 1010, and 1011 CFU/mL (Figure 3a and 3b; linear P<0.01).
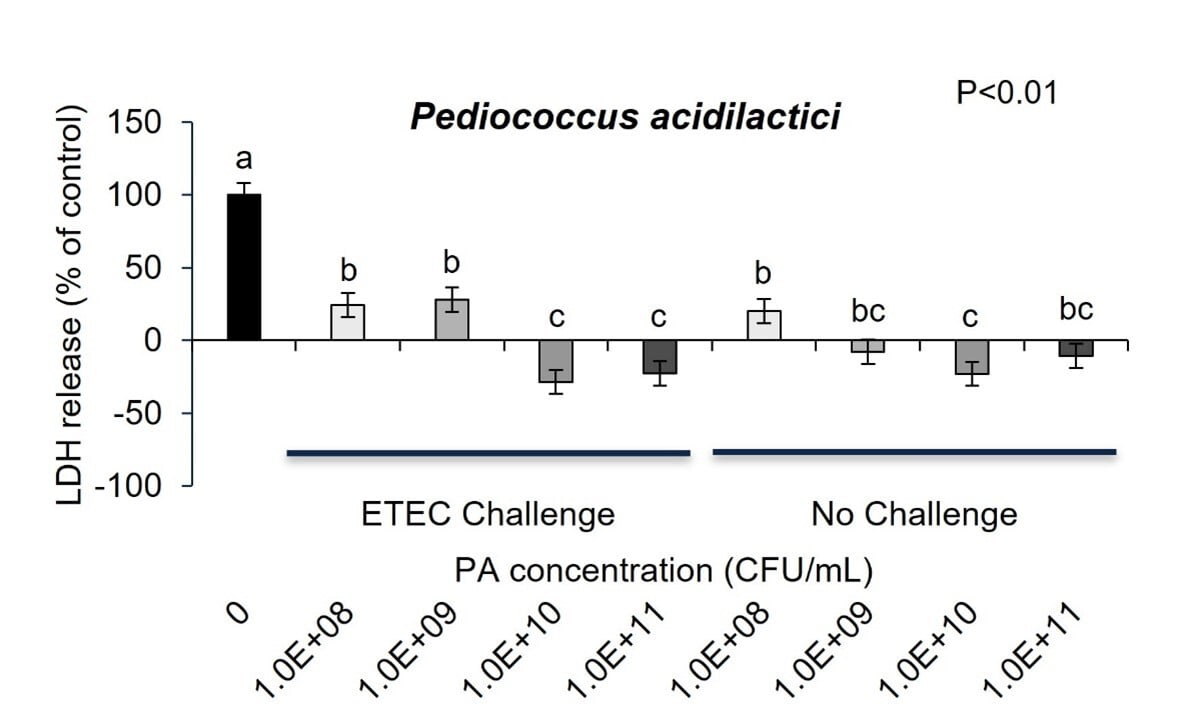
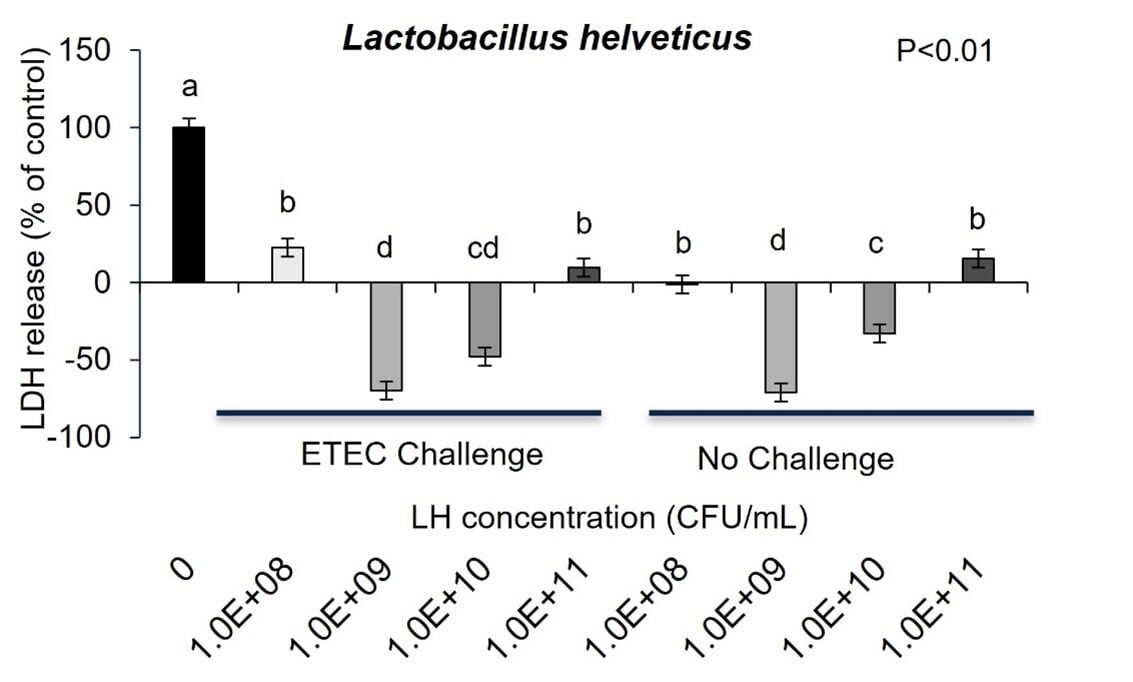
To identify whether ETEC K88 caused intestinal cell membrane damage, LDH in IPEC-J2 cell culture medium was detected. Challenge with ETEC K88 MOI of 2:1 for 3 hours increased LDH secretion from IPEC-J2 cells into culture media indicating increased intestinal cell membrane damage that could affect gut barrier function (Figure 4a and 4b; P<0.01). As expected, P. acidilactici-3A and L. helveticus-4A in co-culture with ETEC inhibited pathogen induced IPEC-J2 cell release of LDH into cell media (Figure 4a and 4b). Further, the probiotics alone cultured with the IPEC-J2 cells had no effect on cell cytotoxicity as measured by LDH release into cell culture media (Figure 4a and 4b) .
Conclusions, future workProbiotics have been used to improve gut health in livestock species. However, probiotic supplementation as a replacement for in-feed antibiotics in livestock does not produce consistent results across animal studies. Given the role of microbial endocrinology on the MGB axis, a deeper understanding of multiple probiotic mechanisms could be beneficial to understand how probiotics affect gut function and the bi-directional communication between gut and the brain.
Results from the probiotic genome sequencing indicate Pediococcus acidilactici-3A and Lactobacillus helveticus-4A both have the genes necessary to synthesize and secrete neurotransmitters associated with regulation of the MGB axis. However, Lactobacillus helveticus-4A did not code for glutamate decarboxylase for synthesis of GABA from L-glutamate. These genomic data provide important information about how this probiotic can communicate with other microbes in the lumen, host epithelial cells, and neuroendocrine circuits. Additionally, both selected probiotics have genes expressed to improve digestibility of lipids and carbohydrates that may be beneficial for neonatal pigs (García-Cano et al., 2019).
These attributes could be useful in supplementation pre-weaning to improve nutrient utilization during weaning stress. Further, data provide evidence in the presence of L-tryptophan there is a dose dependent response of serotonin secretion by Lactobacillus helveticus-4A (Figure 1). However, Pediococcus acidilactici-3A did not increase serotonin secretion in culture with increasing L-tryptophan. This could be related to different applications of these probiotics as the mechanisms are further developed. Research on other Pediococcus probiotic strains show this genus of bacteria is more likely to synthesize and secrete GABA (Anggraini et al., 2019). Preliminary data from our lab would suggest the Pediococcus acidilactici-3A in the presence of glutamate increase GABA secretion (data not shown).
Mechanistically, data indicate Pediococcus acidilactici-3A and Lactobacillus helveticus-4Aboth seem to function in similar manners with adhesion to pig intestinal epithelial cells and inhibition of pathogen, ETEC K88, adhesion (Figure 2 and 3). A fundamental mechanism of probiotics is competitive exclusion of pathogens from the gut environment through multiple mechanisms. Although, it is not fully described if the bacteria in the lumen of the gut or those adhered to the mucosal epithelium have different functions, it would seem logical that probiotic exclusion of pathogen adhesion to the mucosal cell surface would be beneficial to gut health.
In fact, the cell cytotoxicity data supports this conclusion showing that ETEC K88 pathogen induces increased cell membrane permeability while Pediococcus acidilactici-3A and Lactobacillus helveticus-4A did not adversely affect IPEC-J2 cell membrane permeability. Further, when ETEC K88 and Pediococcus acidilactici-3A or Lactobacillus helveticus-4A were co-cultured with the pig intestinal cells the probiotics prevented the ETEC from inducing cell membrane cytotoxicity. Although there is linear dose response in Pediococcus acidilactici-3A and Lactobacillus helveticus-4A from 108 to 1011 CFU/mL in the adhesion of the probiotics to the IPEC-J2 cells, the dose response in inhibition of ETEC adherence or improving cell cytotoxicity did not seem to have as significant of impact on those mechanisms.
Further, in vitro analysis on intestinal barrier function suggest microbiota secretions could have an impact on gut health. We are also working on piglet trials to understand the mechanisms on the gut barrier, microbial endocrine function, and regulation of feed intake centers in the brain. Understanding the mechanisms of these probiotics in a complex environment of the pig intestine may lead to improved consistency of probiotic use in the swine industry to improve production efficiency and animal well-being while providing and an alternative to in-feed antibiotics.
References available upon request
Jacobi is an assistant professor in the Department of Animal Sciences at The Ohio State University.