High dietary levels of zinc oxide modulate the metabolism of trace minerals
Findings may be detrimental to post-weaning piglet health
By Danyel Bueno Dalto, J. Jacques Matte and Jérôme Lapointe
Zinc oxide (ZnO) is commonly used as growth promoters and alternative to antibiotics to prevent diarrhea in weaned piglets. Although the NRC [1] recommended 80-100 mg/kg of dietary zinc (Zn) for 7-25 kg piglets, the swine industry worldwide uses levels up to 3000 mg/kg of Zn during the first weeks post-weaning (Dalto et al., 2020; Yang et al., 2021; Faccin et al., 2023).
Although this nutrient is essential in pig nutrition, the use of such high levels has been challenged due to environmental issues and concerns related to public health (bacterial resistance), which led European countries to restrict its use in swine diets back in 2022. Nevertheless, there is still limited understanding of the interplay between the modulation of Zn in different organs and how it may interfere with the metabolism of other trace minerals. This has hampered advances to replace this strategy without compromising piglets health. For example, high dietary Zn supplementation induces anemia and reduces tissue iron concentrations in rats and chicks (Yanagisawa et al., 2009; Hachisuka et al., 2021). Although the mechanisms controlling this interaction are not well known, some studies indicate an indirect impact of Zn through effects on copper metabolism (Jeng and Chen, 2022). In fact, copper has key roles in iron absorption and utilisation (Arredondo and Núnez, 2005).
A recent trial in our laboratory (Galiot et al., 2018) showed that serum and liver levels of copper decreased (19.6 and 75.6%, respectively) two weeks post-weaning in piglets fed 130 mg/kg of copper. It was hypothesized that this drop in copper status was triggered by the post-weaning high levels of dietary ZnO. Therefore, two studies were designed to better understand the biological consequences of (1) increasing dietary levels of ZnO and (2) different dietary zinc/copper ratios on trace minerals (zinc, copper, and iron) regulation in post-weaned pigs.
One hundred twenty (study 1) and 160 (study 2) piglets were selected at weaning (21 days of age) and randomly assigned to one of experimental post-weaning diets (without antibiotics):
Study 1:Basal diet supplemented with 100, 1000 or 3000 mg/kg of Zn (LZn, MZn and HZn) as ZnO. All three diets had similar supranutritional levels of Cu (130 mg/kg) as copper sulfate (CuSO4).
Study 2:Basal diet supplemented with 100 (LZn) or 3000 (HZn) mg/kg of Zn as ZnO in combination with 6 (LCu) or 130 (HCu) mg/kg of Cu as CuSO4.
Piglets were slaughtered at day 21, 23, 35, and 42 (study 1) and at day 21, 28, and 42 (study 2), for the collection of serum and body tissues.
Growth performanceThe use of supranutritional levels of dietary ZnO in post-weaning diets is known to promote growth performance although some studies have not detected such beneficial effects (Espinosa et al., 2020ab). In both present studies, high levels of dietary Zn impaired growth. Particularly in study 2, the combination of high levels of dietary ZnO and CuSO4 (HZnHCu) impaired piglets growth whereas beneficial effects of high dietary CuSO4 levels were detected when low ZnO levels were used. The optimized experimental environmental conditions of these studies may have interfered with the mainly local (intestinal lumen) effects of ZnO. Under these conditions, it is possible that the negative effects of high ZnO levels on metabolism (see below) have overweighed its known positive effects observed under more challenging environments (high pathogenic pressure). For CuSO4 that has mainly systemic actions, the influence of the experimental environment was likely less disturbing.
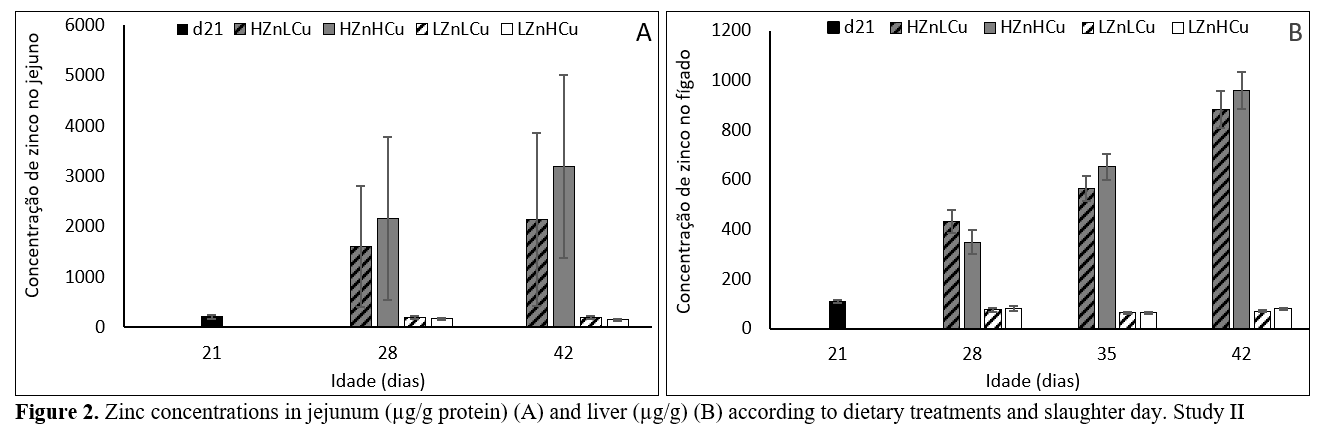
ZincIn both studies, although HZn diets decreased the expression of genes related to intestinal Zn absorption, a proportional reduction in jejunum, liver, and serum Zn concentrations (Figure 1A, 2A and 2B) was not observed. To avoid Zn toxicity, different cellular mechanisms were activated in jejunum and liver aiming to (1) trap Zn within cells and (2) release excess Zn to the systemic circulation. Despite the activation of these regulatory mechanisms, sharp increases (approximately 5- and 12-fold in serum and liver, respectively) were detected in HZn groups of both studies during the three weeks of supplementation compared to pre-treatment values (d21).
In study 1, the 10-fold difference in dietary ZnO levels between LZn and MZn resulted in approximately 2-fold increase in serum levels, indicating modulation of Zn concentrations. However, the 3-fold difference in dietary ZnO levels between MZn and HZn resulted in a proportional increase in serum Zn levels, suggesting that somewhere between 1000 and 3000 mg ZnO/kg of diet the mechanisms of regulation were no longer efficient in managing the excess of Zn.
Considering both studies, serum Zn concentrations in piglets supplemented with 3000 mg ZnO/kg of diet ranged between 4.1 and 4.6 mg/L at d42, levels that were reported to impaired performance in pigs (Hahn and Baker, 1993), which is in line with the lower growth performance in those animals.
In study 2, liver Zn concentrations in LZn groups at d28, 35, and 42 represented 71, 58, and 66%, respectively, of pre-treatment values (d21) whereas serum values remained constant throughout the experimental period. Such results suggest that 100 mg ZnO/kg of diet during the first weeks post-weaning may not completely fulfil the piglets needs.
CopperIndependently of dietary Cu levels (130 ppm in study 1 and 6 or 130 ppm in study 2), piglets in HZn groups had greater jejunum Cu concentrations than those in LZn (Figure 3A), which was concomitant with lower liver and serum Cu concentrations (Figure 1B and 3B). These results suggest that dietary Cu was absorbed by intestinal cells but was not further released to reach the liver.
This impairment of Cu efflux from intestinal cells is related to the activation of a mechanism responsible for trapping minerals in different organs. This mechanism is controlled by an enzyme (metallothionein; MT) that is mainly responsive to dietary Zn levels but that has a much greater affinity for Cu. In study 1, this enzyme was 357- and 250-fold more expressed in jejunum and liver, respectively, of HZn compared to LZn piglets three weeks post-weaning. The high expression of MT in liver of piglets fed HZn, that would promote trapping of Cu in hepatic cells, was expected to increase liver Cu concentrations but, in contrast, values were not far from those detected in LCu groups (Figure 3B), which likely did not receive enough dietary Cu (see below).
These counter intuitive results are explained by the fact that the high hepatic Zn content stimulates the expression of MT in liver, and that irrespectively of the low hepatic Cu concentrations, which were limited either by the low Cu intake (LCu; study 2) or by the significant sequestration of Cu within intestinal cells by MTs (HZn in study 1 and 2). The greatest hepatic and serum concentrations of Cu in LZnHCu groups in study 2 further support this reasoning. Considering the fast renewal of the intestinal tissue (25% per day), this trapping of Cu in intestinal cells may work as mechanism of Cu excretion because it provides a transient storage for this mineral that is eventually lost through slough.
Interestingly, in study 2, hepatic Cu concentrations were lower than pre-treatment values (d21) from d28 not only for HZn but also for LCu groups (Figure 3B) whereas serum values (Figure 1B) were lower for all treatments. Besides further supporting that HZn diets impair Cu metabolism independently of dietary Cu levels, it also indicates that 6 mg of Cu/kg of diet (LCu, study 2) may not be sufficient to fulfil the post-weaning piglets requirements for this mineral.
IronHigh dietary ZnO levels impair Fe metabolism by systemic and local (intestine and liver) mechanisms. In both studies, piglets supplemented with HZn had an apparent impairment in the intestinal absorption of Fe which was concomitant with an likely increased sequestration of Fe in enterocytes, resulting in reduced hepatic Fe content. High dietary ZnO levels have also stimulated a greater release of iron from hepatic cells, which may have exacerbated the low storage of iron in liver. However, these effects were not intense enough to impact hemoglobin, whole blood and serum iron concentrations. Therefore, high dietary levels of ZnO in post-weaning diets may not cause iron deficiency in post-weaning piglets but interfere with hepatic Fe storage, which may be problematic considering the increasing detection of piglets weaned with low iron reserves in recent years (Bhattarai and Nielsen, 2015; Perri et al., 2016; Almond et al., 2017; Gillespie, 2019).
This last point is very important because observed differences in iron metabolism between both studies suggest that pre-weaning factors impacting Fe metabolism may interfere with the post-weaning homeostasis of this trace mineral, which deserve to be further investigated.
Conclusions During the first weeks post-weaning, piglets zinc requirements appears not to be satisfied by 100 mg of zinc as ZnO/kg of diet. On the other hand, high dietary levels of ZnO were not efficiently modulated in different organs resulting in remarkably high serum zinc concentrations that may likely have reduced piglets growth performance, besides impairing copper and iron metabolism.
The low concentrations of copper in liver and serum indicate the potential risk of copper deficiency in piglets supplemented with high levels of dietary ZnO during the post-weaning period. However, independently of these major zinc effects, a level of 6 mg of copper/kg of diet apparently did not meet the copper requirements of post-weaning piglets. Regarding iron, the present results suggest that supplementation with high levels of ZnO interfere with the storage of iron in liver but would not induce iron deficiency in post-weaning piglets.
Although the use of lower levels of dietary ZnO are recommended to avoid such detrimental effects on piglets health (in addition to those on environmental and public health), there is still no consensus regarding the most appropriate levels of Zn supplementation to post-weaning piglets.
Dalto and LaPointe are research scientists and Matte is an honorary scientist, all with Agriculture and Agri-Food Canada.