Dietary strategies to improve pig health should be based on goals of the production unit
By Dan Columbus
A number of factors contribute to the gap observed between performance potential and actual performance achieved under commercial conditions, with immune status likely playing a significant role. Pigs are continuously exposed to microbial pathogens and immune-stimulatory antigens that negatively impact animal productivity. This decrease in performance can have a substantial impact on profitability of producers.
Impact of disease challenge on performance and nutrient metabolismA primary result of immune challenge is a reduction in feed intake, with subsequent reduction in growth performance of pigs. While feed intake generally explains a majority of this reduced growth, recent meta-analyses have demonstrated that both reduction in nutrient supply (i.e., feed intake) and nutrient utilization (i.e., maintenance requirements) are contributing factors. The proportion with which these factors impact performance is dependent on the specific challenge type, with reduced growth during enteric bacterial infections being more a result of changes in nutrient utilization while the negative effects of mycotoxins and respiratory disease are more due to decreased feed intake (Pastorelli et al., 2012; Rodrigues et al., 2021a).
The overall magnitude of response and recovery period is also dependent on the sex and age at which the pig is exposed to the immune challenge. Rodrigues et al. (2021a) found that female pigs showed a greater negative response in performance compared to males and that the reduction in male pigs appears to be more related to alterations in nutrient utilization than the reduction in female pigs. It was also shown that younger pigs (e.g., nursery) are more affected by immune challenge than older pigs, however, older pigs take longer to recover to pre-challenge performance levels.
Regardless of the type of immune challenge, stimulation of the immune system alters protein and amino acid metabolism and utilization, with amino acids redirected from growth towards supporting the immune response. As feed intake also decreases during immune stimulation, the increase in amino acid supply to support the immune response is met, in part, through a reduction in protein synthesis and increase in protein catabolism in the muscle, which represents the largest pool of amino acids. However, the amino acid profile of muscle protein differs significantly from that of protein involved in the immune response (Reeds et al., 1994), resulting in an amino acid imbalance and a disproportionate use of some amino acids during immune challenge and obligatorily lead to an increase in whole-body amino acid catabolism and reduction in body protein growth. For example, based on amino acid profiles of skeletal muscle protein and acute phase proteins, 6 g of muscle protein would be required to meet the demands for cysteine for every 1 g of albumin (Rakhshandeh, 2011).
Overall, while it is true that immune challenge will result in a reduction in feed intake and, therefore, reduce growth and nutrient requirements for growth, it is likely that amino acid requirements (i.e., amino acid ratios to lysine) need to be adjusted during times of immune challenge and requirements determined in healthy pigs are, most likely, inappropriate.
Nutrient effects on gut health and immunityA number of dietary interventions have been assessed for their potential to improve gut health and overall health in pigs and as potential replacements to antibiotics, including organic acids, phytochemicals, enzymes, and probiotics, and many reviews have been published on their use in swine. Here, the focus will be on dietary nutrient content, including dietary protein, fibre, and amino acids and their role in promoting health in the pig.
Dietary protein and fibre content can have significant impacts on both animal performance and health, largely through effects within the gastrointestinal tract (Jha and Berrocoso, 2016). The impact of protein and fibre on animal health is increasingly important to understand with the increased use of co-products and other novel feedstuffs which generally have a greater fibre content and variable protein content and protein availability (Pieper et al., 2012a; Zijlstra and Beltranena, 2013), especially when such ingredients have undergone heat processing.
Dietary protein, while necessary to meet requirements for growth in pigs, can have a detrimental effect on gastrointestinal health if provided in excessive amounts. Dietary indigestible protein content (i.e., protein not absorbed in the small intestine) is available for microbial fermentation and may have detrimental effects on gut health. Metabolites of protein fermentation, including branched-chain fatty acids, ammonia, biogenic amines, hydrogen sulfide, and phenolic and indolic compounds (Pieper et al., 2012a; Yao et al., 2015; Jha and Berrocoso 2016)), have been associated with toxic and pro-inflammatory effects on the gut epithelium, including compromised colonic epithelial cell structure and metabolic functions, thinning of the mucus barrier, and increased colonic permeability (Gaskins, 2001; Hughes et al., 2008; Yao et al., 2015).
Therefore, it has been suggested that high protein diets increase susceptibility to enteric pathogens and are a predisposing factor in the development of post-weaning diarrhea. Further, it has been suggested that feeding a low protein diet will reduce the amount of substrate available for the proliferation of pathogenic bacteria and thus minimizes production of associated toxic metabolites, which in turn will improve gut health and function in piglets (Nyachoti et al., 2006).
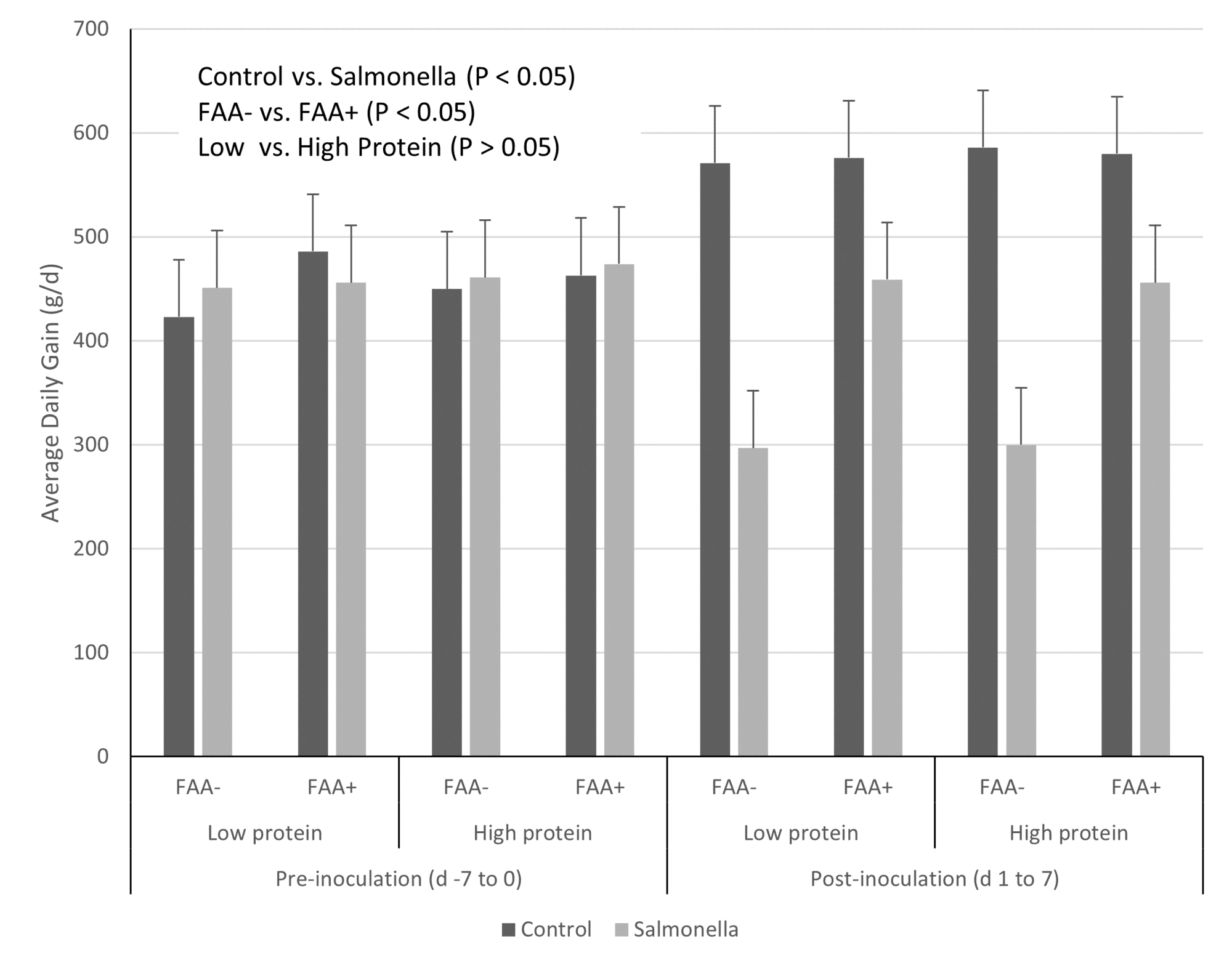
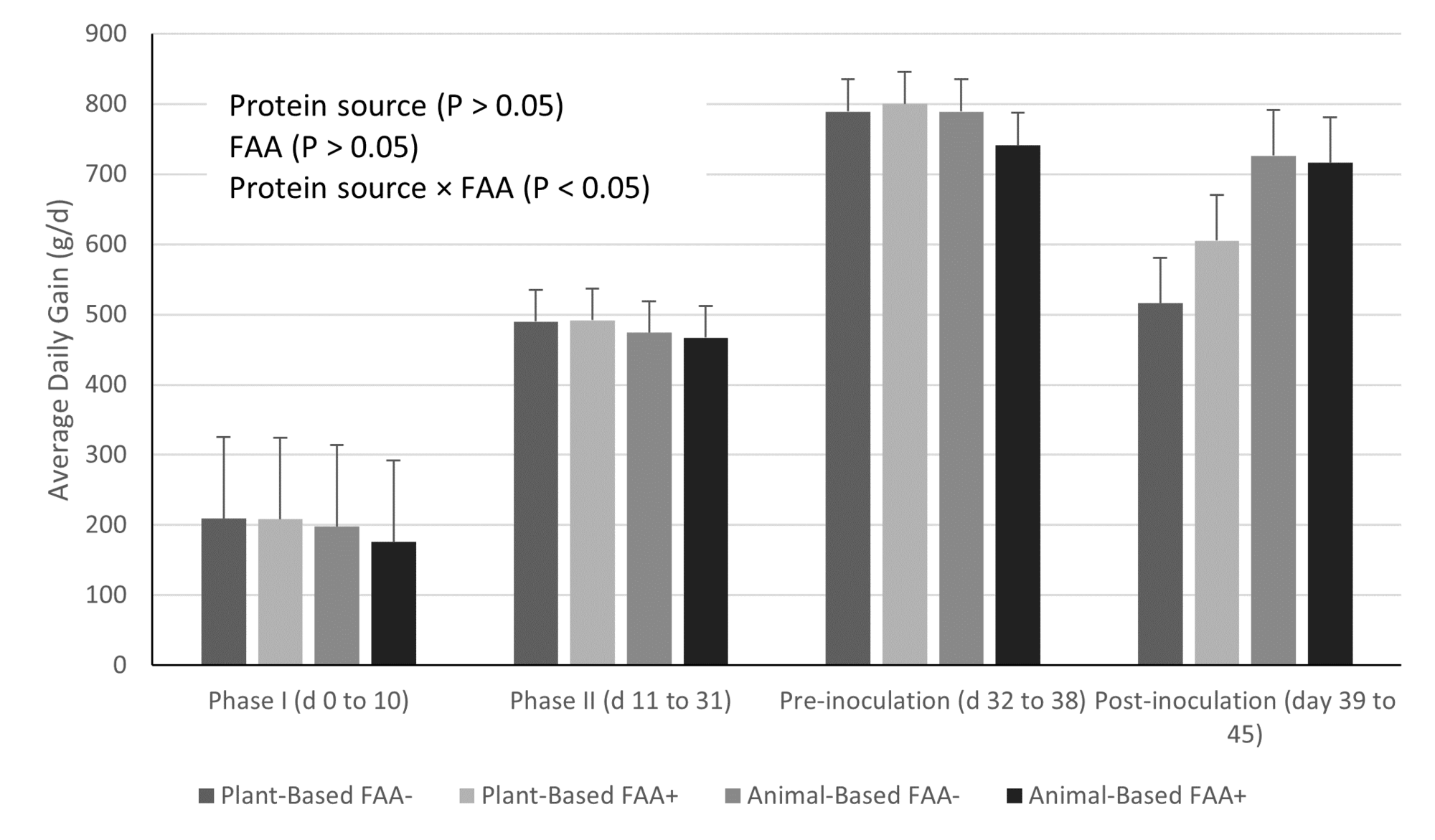
However, while there is a general trend for reduced incidence of diarrhea with decreasing dietary protein content, this is not consistent across studies when examining the same protein content, suggesting a factor other than simply total dietary protein content (e.g., indigestible content, protein type) is involved. For example, while increased dietary protein content had no impact on pig response to a Salmonella challenge in one study (Fig. 1; Rodrigues et al., 2021b), there was a significant difference in response to a subsequent challenge when pigs were fed either plant-based or animal-based protein sources in nursery diets (Fig. 2; Rodrigues et al., 2022a).
This suggests that digestibility, which is greater in animal-based products, anti-nutritional factors, which are greater in plant-based products, and/or bioactive compounds present in animal-based products contribute to the observed response to dietary protein content and post-weaning diarrhea and susceptibility to enteric pathogens.
Unlike with protein, fermentation of fibre is generally considered to result in production of beneficial metabolites, such as short-chain fatty acids, that promote gut health and limit pathogen growth (Pieper et al., 2012ab; Jeaurond et al., 2008). As with protein, the impact of dietary inclusion of fibre on nursery pig performance and intestinal health has been inconsistent, likely due to differences in the physicochemical properties and fermentability of different fibre sources. Inclusion of a non-structural/soluble fibre source may provide intestinal bacteria an alternative substrate for fermentation (Pieper et al, 2012ab) whereas inclusion of structural/insoluble source of dietary fibre may reduce the impact of indigestible protein through increased digesta flow through the gut and reduced adhesion of pathogens (Molist, 2020).
Two studies examining the impact of a blend of soluble and insoluble fibres (i.e., wheat bran and sugar beet pulp) demonstrated the potential benefits, but also the variability in response, of dietary fibre inclusion to reduce effects of high indigestible protein content. Pieper et al. (2012a) showed that while fermentable protein increased production of harmful metabolites and altered the mucosal response in weaned pigs, this was not impacted by inclusion of fibre, although higher fibre inclusion did beneficially impact the microbial environment.
Alternately, in grower pigs, Wellington et al. (2020a) demonstrated a negative impact of indigestible protein content on metabolite production and mucosal response, which was partially attenuated (i.e., improved short-chain fatty acid/branched-chain fatty acid production and intestinal barrier gene expression) with inclusion of the fibre blend (i.e., sugar beet pulp and wheat bran). The benefit of dietary fibre inclusion on gut health has also been demonstrated by Wellington et al. (2020b), who observed improved barrier function in pigs challenged with either E. coli lipopolysaccharide or Salmonella through increased mucin production and production capacity (i.e., goblet cell number).
Functional amino acids and pig healthOf the amino acids, glutamine, arginine, threonine, and aromatic and sulfur amino acids are of particular importance during immune challenge (Reeds and Jahoor, 2001) and provision of these amino acids in excess of requirements for growth during immune challenge may be of benefit in mitigating the effect of immune challenge on growth performance.
Previous studies have demonstrated an increased requirement for growth for methionine and cysteine (Rakhshandeh et al., 2010; Litvak et al., 2013), threonine (Jayaraman et al., 2015; Wellington et al., 2018; McGilvray et al., 2019), and tryptophan (De Ridder et al., 2012) in response to immune stimulation.
As indicated above, while past research has indicated alterations in nutrient metabolism, generally, and amino acid requirements, specifically, only recently has the effect of supplementation with amino acids for pig robustness received significant attention.
The term ‘functional amino acids’ has been used to describe those amino acids supplemented in the diet for their roles beyond those for protein synthesis (i.e., lean gain). In the context of health, these include amino acids with significant roles in gastrointestinal health (e.g., barrier function) and immune status (e.g., antioxidant balance, acute phase response).
Supplementation of individual functional amino acids has been shown to improve growth performance of pigs under immune challenge. For example, Wellington et al. (2019) showed that supplementation with threonine resulted in improved growth performance of growing pigs fed high fibre diets during Salmonella challenge most likely through support of improved mucin production, as indicated by increased fecal mucin output.
Likewise, Koo et al. (2020) observed improved gut integrity (i.e., goblet cell density, tight junction gene expression) in pigs fed a ‘simple’ diet which induced an intestinal inflammatory response.
Jayaraman et al. (2015) and Trevisi et al. (2015) observed improved growth performance in pigs housed in unsanitary conditions and challenged with E. coli, respectively, when fed diets containing supplemental threonine. In another study, Koo et al. (2021) observed improved immune status in pigs fed supplemental valine and Jayaraman et al. (2015) and Le Floc’h et al. (2009) observed improved growth performance in weaned pigs provided supplemental tryptophan when housed in unsanitary conditions.
Recent work has examined the use of a blend of functional amino acids on growth performance and immune status in weaned pigs. Rodrigues et al. (2021b) fed diets containing either a standard amino acid profile (NRC, 2012) or one in which threonine, methionine, and tryptophan were supplemented at 120% of the requirements for growth. When challenged with Salmonella, those pigs that had been provided the supplemented amino acid profile had improved growth performance and immune status (i.e., acute phase response, antioxidant balance) than those that had received the basal profile.
Supplemental amino acids also resulted in a decrease in fecal myeloperoxidase, an indicator of intestinal damage, indicating a role of these amino acids in supporting gut health during an enteric pathogen challenge.
The effectiveness of supplementation with this amino acid blend may be dependent on specific conditions, as van der Meer et al. (2016) showed improved immune status but limited effect on growth performance from wean to finish when pigs were given the same amino acid blend and supplementation level but under low or high sanitary conditions.
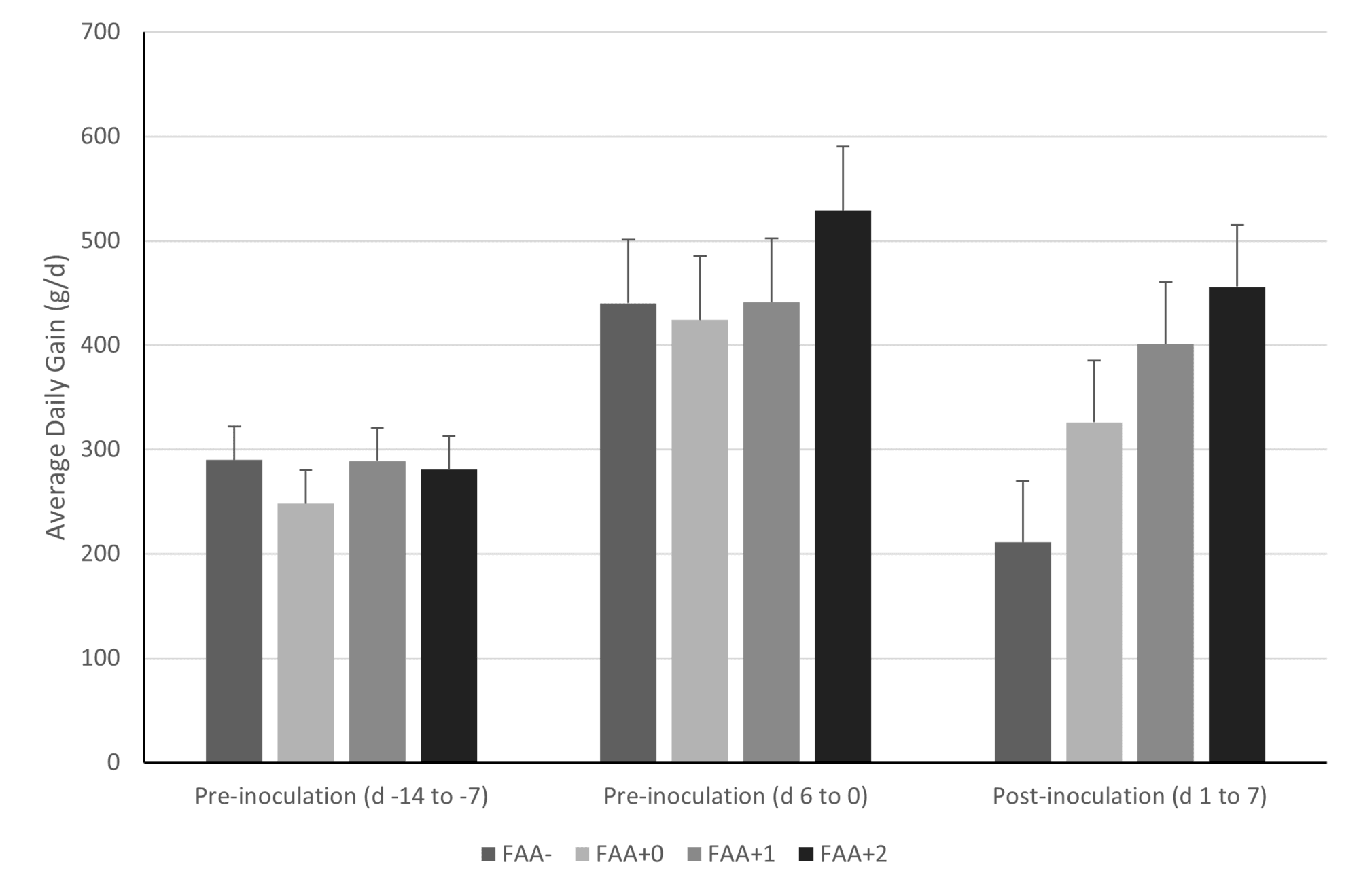
The effectiveness of dietary interventions for improved pig performance during health challenge may be dependent on a number of factors. The timing of functional amino acid supplementation may be important with respect to their effectiveness at improving performance and immune status. For example, Rodrigues et al. (2021c) demonstrated a further increase in growth performance in pigs provided the blend of functional amino acids when the adaptation period prior to Salmonella infection was increased from 0 to 2 weeks (Fig. 3).
However, the response to functional amino acids was variable with respect to immune status in these pigs, with acute phase response responding positively to a longer adaptation time while antioxidant balance did not.
While there was no overall effect of amino acid supplementation, van der Meer et al. (2016) observed improved average daily gain in the nursery period and improved feed efficiency in the finisher period in pigs housed in unsanitary conditions. Moreover, the response to sanitary conditions was greater during the nursery period, indicating that supplementation may be more beneficial in the immediate post-weaned period.
Schweer et al. (2019) and Jasper et al. (2020a) were able to improve growth performance in pigs inoculated with PRRS virus by adjusting overall dietary nutrient content (i.e., increasing lysine:energy) at the time of challenge, however, no improvement was observed if the adjusted diet was provided post-challenge (Gabler, 2021).
Functional amino acids may provide more long-term benefits, as Rodrigues et al. (2022a) demonstrated improved growth in pigs that had received a supplemental functional amino acid blend in the nursery period but were fed a common grower diet at the time of Salmonella challenge. This blend of amino acids, however, was only partially able to attenuate the negative effect of disease challenge in low-birth weight pigs during the subsequent disease challenge.
Other components in the diet may also affect functional amino acid effectiveness, as supplementation with the same blend of functional amino acids was able to improve performance of nursery pigs that had received a plant-based diet, however, provided no further benefit to those pigs that had received animal-based protein sources in the nursery (Fig. 2; Rodrigues et al., 2022b).
The appropriate intervention may be dependent on the type of challenge experienced, and is likely related to the specific immune response to different challenge types and the proportion of the decrease in performance that is due to either reduced feed intake or alterations in nutrient utilization (Rodrigues et al., 2021a, Pastorelli et al., 2012).
For example, while nutrient adjustment was effective in PRRS infected pigs, this adjustment was not effective in pigs challenged with Mycoplasma hyopneumoniae (Jasper et al., 2020b). This may also explain why the blend of functional amino acids was effective during an enteric pathogen challenge (Rodrigues et al., 2021bc) but had reduced effectiveness during a sanitary challenge (van der Meer et al., 2016).
ConclusionsDisease challenge can have a substantial negative effect on productivity and profitability of swine production, however, there are a number of dietary strategies that can be used to improve pig robustness.
Dietary protein and fibre can be adjusted to promote a healthy gut environment. Recently, functional amino acids have been investigated for their role during disease challenge and ability to support both the immune response and growth performance. While these have been shown to be effective under some circumstances (i.e., normal birth weight, enteric challenge), this is not always the case.
Overall, dietary nutrient content is another tool that can be incorporated into pig health programs. Dietary strategies to improve pig health should, however, be based on the specific circumstances and goals of the production unit.
References available upon request
Columbus is a research scientist in nutrition at Prairie Swine Centre.