Soybean meal mitigates respiratory disease–impaired growth in pigs
This is the first of 5 articles based on the presentation ‘Soybean meal: Growth and health-promoting effects during high health and immune stress’ at the John F. Patience, Int. Swine Nutrition Conf. (August 2021) at Iowa State University
By R. Dean Boyd, Mike Johnston, James Usry, Paul Yeske and Aaron Gaines
Introduction
Soybean meal (SBM) is a superb amino acid and energy source for pig diets, but another important dimension is emerging in relation to health-promotion in humans and food animals. Soybeans contain an abundance of functional molecules (FM) in the non-nutritive fraction (Sharma et al., 2019), as well as bioactive peptides that arise during digestion of major SBM proteins (Chatterjee et al., 2018). There is considerable information on these in relation to human health, but research with food animals is in the early stages of development (Smith and Dilger, 2018).
To our knowledge, the first illustration of the disease-mitigating effects of SBM in pigs was reported in 2010 by researchers at the Hanor Company (Johnston et al., 2010; Boyd et al., 2010). Swine respiratory disease infection (SRD) was unexpectedly encountered during a study with growing pigs that involved low SBM (typical) and high SBM diets. Pigs fed low SBM diets exhibited a profound loss in growth and feed conversion efficiency (FCE), which is common during periods of high immune stress. However, pigs fed diets with higher SBM content (38-50%) grew as though no active infection was present. These results were confirmed by another group who conducted a similar study design, almost simultaneous with ours, and who also encountered SRD pathogens unexpectedly (Gene Gourley, M.S., personal communication).
In effect, the planned experiment at Hanor became decidedly more valuable when pigs encountered a SRD complex of pathogens. This proved to be fortuitous. Our observation that SBM could mitigate SRD–impaired growth was novel for food animals and led to a new frontier in ingredient science.
This paper describes that work and makes the case for a more balanced use of SBM as compared to extreme displacement by alternative amino acid sources during periods of SRD stress. Results from the Hanor study exposed the hidden cost of SRD and illustrated the effectiveness of SBM in mitigating impaired growth. Our observation that SBM can mitigate the effects of SRD is strengthened by results from a commercial study where the ability of pigs’ to thrive improved by increasing dietary SBM content (Cemin et al., 2020). We conclude with the proposition that SBM is both a nutritional and health-promoting ingredient.
Intent of the original Hanor study
The original experiment was designed to determine whether SBM level was a factor in meeting the dietary lysine requirement for pigs fed the metabolic modifier, ractopamine hydrochloride (RAC). In other words, did the level of dietary lysine required to maximize growth and FCE depend on the amount of SBM used to meet the lysine requirement? The simultaneous investigation of response to dietary lysine and SBM level was a fortunate design feature when SRD infection was unexpectedly encountered. Given this circumstance, the question evolved beyond how amino acid specifications were met to whether the nutraceutical components of SBM could mitigate respiratory disease impaired-growth. In the end, the high SBM (H-SBM) regimen profoundly improved the level of growth and FCE achieved when compared to low SBM (L-SBM) fed pigs.
Study design
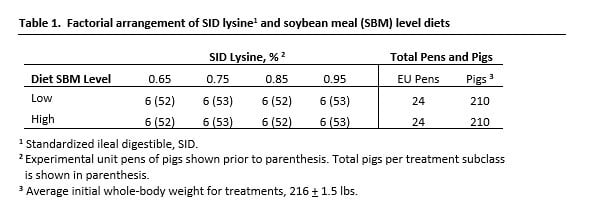
Dietary treatments were arranged as a 4 x 2 factorial; having 4 standardized ileal digestible (SID) lysine levels and 2 SBM levels (L-SBM, H-SBM). Pigs in all 8 diet subclasses were fed RAC (5 ppm) with whole-body growth rate (ADG), FCE and carcass gain being the key measures. A total of 420 castrate pigs (PIC terminal genetics) with an average body weight of 217 lbs were used. Data were collected each week for 21 d, after which all pigs were harvested. The commercial research facility that Hanor used was an academic equivalent setting, since pig density (room, pen) was low and environmental stressors normally encountered in commercial sites were reduced. The latter was accurate until we encountered an active infection of SRD complex of pathogens. The experimental framework is shown in Table 1.
Diet formula design
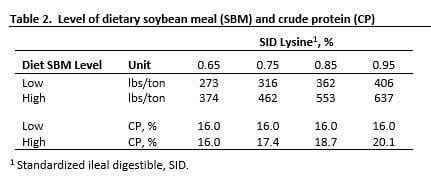
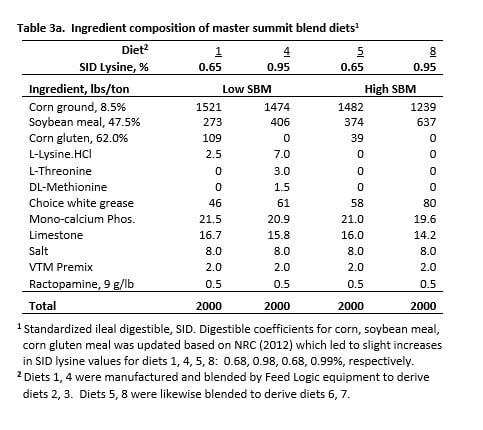
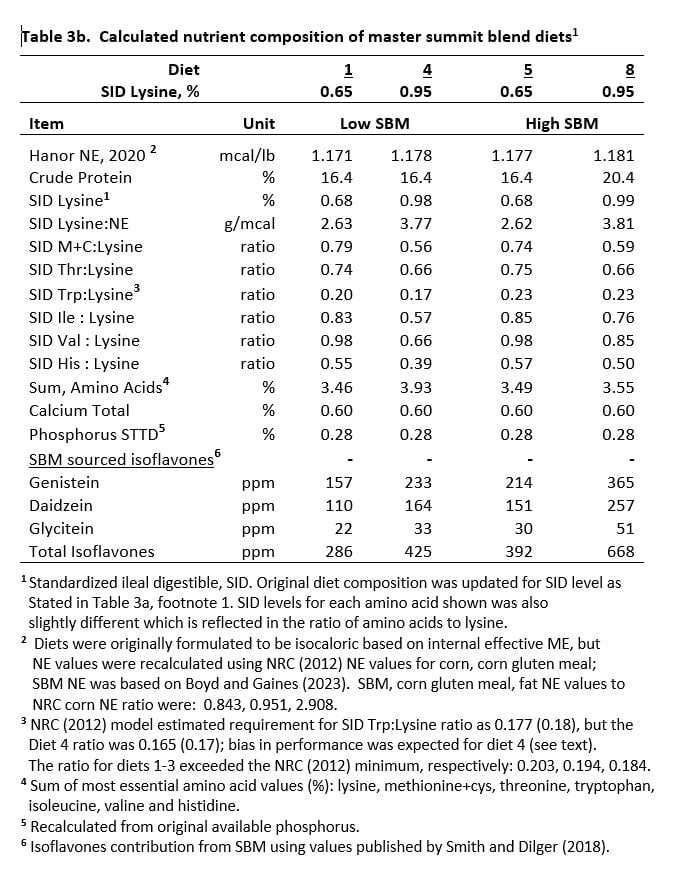
Diets were formulated to provide 0.65, 0.75, 0.85 or 0.95% SID lysine for both SBM regimens. The level of SBM and crude protein (CP) for each diet subclass is shown in Table 2. Diet composition is shown in Table 3a with calculated nutrient content for diets shown in Table 3b. Nutrient level was confirmed by analysis.
Four diets were manufactured and summit blended at the research farm to create the 8 experimental diets using Feed Logic equipment. Diets 1 and 4 were composed for L-SBM diets with SID lysine levels achieved by increasing SBM and crystalline amino acids. Diet 1 (L-SBM) included corn gluten meal to meet the minimum CP level (16.0%) required for diets containing RAC. Diets 5 and 8 were manufactured for H-SBM diets by increasing SBM content, without the use of crystalline amino acids. This allowed comparison of the growth response to dietary lysine for typical diets (L-SBM) to those with significantly higher SBM content.
The 0.65% SID lysine diet for both SBM levels (diets 1, 5) was equivalent for CP (16.0%) and for each of the 6 most limiting essential amino acids and histidine; their total being 3.46, 3.48% for L-SBM and H-SBM diets, respectively. Both diets were isocaloric (net energy, NE). Nutrient equality was important because a difference in response at that point (0.65% SID lysine) could not be attributed to different levels of CP, essential amino acids or NE. Given the extreme care to achieve nutrient equivalency, this diet pair was the anchor for the response curve with respect to the interpretation of results.
The remaining test diets (0.75, 0.85, 0.95% SID lysine) were equivalent for NE, minerals and vitamins, but not for all essential amino acids or CP. However, each diet met or exceeded the requirement for the 5 most limiting essential amino acids in relation to dietary lysine, based on the current NRC standard (2012). The CP level for each lysine cohort pair was higher for H-SBM diets (Table 2), which led to greater amounts of isoleucine, valine and dispensable amino acids. Diets were formulated based on nutrient specifications from the literature at that time, but amino acid and NE levels were recalculated (Table 3b) and evaluated based on the 2012 NRC standard for this publication.
Fortunately, estimates for the SBM NE to corn NE ratio that was used for this experiment were in relative agreement with recent estimates for academic-equivalent settings (Boyd and Gaines, 2023). The updated estimates for SBM and corn are reflected in calculated NE values shown in Table 3b.
Unexpected infection with respiratory pathogens
Pigs unexpectedly became infected with multiple respiratory pathogens (SRD complex) that triggered systemic inflammation. SRD causes both growth and FCE to plummet compared to unaffected pigs. Upon learning of the infection, all pigs received medication by water and clinical diagnostics and physical inspection of pigs was initiated under the direction of Tara Donovan, DVM (The Hanor Co.). Serology revealed that pigs were PRRSv and porcine circovirus (PCV2) positive. There were PCV2 lesions and Streptococcus suis infection was also confirmed. The inflammatory nature of these viruses has been proven by the presence of circulating pro-inflammatory cytokines (Parra et al., 2006).
The study continued as planned because SRD infection is a production reality. In North America, SRD complex is prevalent in many systems during January – April when barns are closed due to cold weather. In addition, SRD is often a problem for weaned pigs that are derived from PRRSv active sow farms. Taken together, SRD infection can span 6-7 months of the year.
SBM mitigated respiratory disease-impaired growth
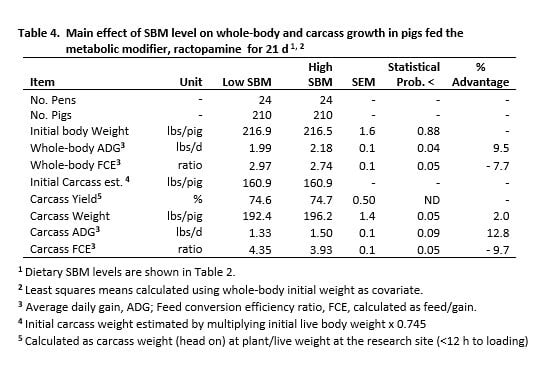
The overall effect of SBM level on whole-body and carcass growth and FCE is shown in Table 4. Since the response to dietary lysine level was similar for both SBM levels (SBM x lysine level, P>0.29, all criteria), the average response to SBM level is shown for the 21 d study. The growth suppressing effects of SRD is illustrated by comparing the response of L-SBM and H-SBM diets; the latter resulted in profound improvements in whole-body (ADG, +9.5%; FCE, -7.7%) and carcass growth (ADG, +12.8%; FCE, -9.7%).
The net result of H-SBM diets, during the infection, was improved carcass weight gain (3.8 lbs/pig) and significant feed savings. If H-SBM fed pigs were harvested on a weight constant basis to L-SBM pigs (31.5 lbs carcass gain/pig), they would have required 13.2 lbs less feed per pig.
This was the first study to show that SBM can reduce high immune stress-impaired growth. Until this report (Johnston et al., 2010), efforts to reduce disease impaired-growth by nutritional means had been unremarkable. We propose that SBM is a nutritional and ‘prescriptive’ ingredient when SRD is encountered in commercial practice; prescriptive, meaning that SBM is a credible, tactical option for respiratory disease mitigation in pigs (term first suggested by Dr. Lisa Weaver, personal communication).
Growth response was dependent on SBM level
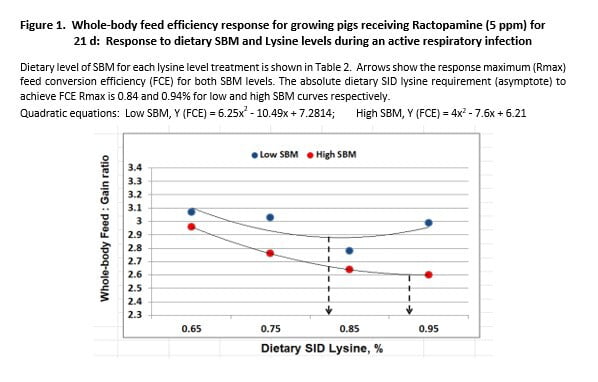
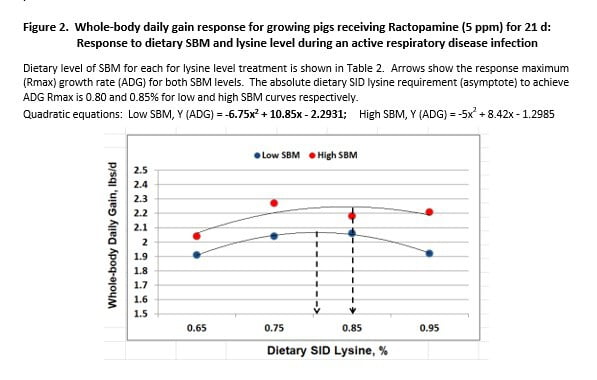
Although the shape of FCE response curves was similar for both SBM regimens, dietary SBM level caused the curves to separate into 2 planes of genetic expression (Figure 1). The same pattern was observed for growth rate (Figure 2). This suggests an important principle in relation to genetic expression when confronted with disease. SBM content influenced the level of growth permitted, with growth depending next on amino acid intake. To illustrate, when higher lysine levels (amino acids) were fed to improve performance by apparently less healthy pigs (L-SBM), FCE and ADG became worse not better (Figures 1, 2).
In the hierarchy of growth regulation, amino acid intake is important but secondary to health status. Physiologically, the body responds to systemic immune stress by prioritizing nutrients for life-sustaining processes over growth (e.g., IGF-1 declines; Johnston, 1997; Spurlock, 1997). Ultimately, amino acid intake provides the building blocks for muscle growth that health status permits.
This research provides an answer to the often asked question, ‘can anything be done nutritionally to improve growth’ when disease is encountered. Diet SBM content, rather than elevated amino acid (or energy) intake, is a means of profoundly mitigating growth suppression by the SRD complex.
SBM level differences were compared at the point of response maximum for FCE and growth. Pigs fed H-SBM diets achieved a minimum FCE of 2.60, compared to 2.88 for L-SBM fed pigs; a difference of 0.28 units (Figure 1). The dietary lysine level required to achieve minimum response also differed. We estimate that 0.94% SID lysine was required for H-SBM fed pigs and that 0.84% was required for L-SBM pigs (asymptote estimates). The 0.28 unit spread in FCE is a key measure in estimating feed cost for unchecked, respiratory disease stress.
Impressive differences were also observed for maximum ADG. H-SBM fed pigs gained a maximum 2.24 lbs/d compared to 2.07 for L-SBM pigs (0.17 lbs/d change, Figure 2). The SID lysine level required to achieve respective maximums was 0.85% for H-SBM fed pigs compared to 0.80% for their counterparts.
Is SBM mitigation of SRD effects nutritional?
The biological basis for the mitigating effects of SBM on SRD impaired-growth is unclear, but it is unlikely to be nutritional. This position is based on NE and nutrient equivalency of cohort diets that anchor (0.65% SID lysine) the growth response curves (Figures 1, 2). SBM level is the variable in play and the difference in SBM FM intake is markedly different. These components are most likely involved in mitigating mechanisms.
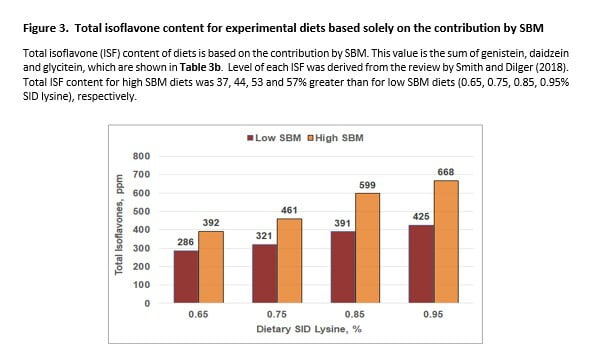
Functional molecules in SBM are remarkably diverse and abundant. The isoflavone (ISF) class of FM was used as a ‘marker’ to track relative FM differences between SBM levels over the lysine response curve. The H-SBM anchor diet (0.65% SID lysine) provided 37% more ISF than the L-SBM counterpart (Table 3b; diets 1, 5). The disparity in ISF content between diet cohorts increased as lysine level increased (Figure 3).
The performance advantage of H-SBM fed pigs was compared across the lysine response curve. Regression-predicted means for FCE and growth response for anchor diets exhibited a 0.14 FCE unit advantage for H-SBM pigs (Figure 1). Growth rate was improved by 0.15 lbs/d over L-SBM counterparts (Figure 2). These differences are nearly twice their respective SEM (Table 4), despite being nutritionally equivalent.
Response variance for SBM level was also compared for 0.75 and 0.85% SID lysine cohort diets. The relative advantage for H-SBM fed pigs at each lysine level was similar to the difference for anchor diets, with some exception. Whole-body FCE differed by -0.15 and -0.25 units (Figure 1) and growth rate improved by 0.15 and 0.19 lbs/d for 0.75% and 0.85% SID lysine diets, respectively (Figure 2).
Comparisons were not made at the 0.95% level because it exceeded the lysine requirement of L-SBM for FCE and ADG. In addition, the L-SBM diet appeared to be slightly deficient in tryptophan based on the NRC 2012 standard (Table 3b), which could introduce nutrition bias into the response. Each of the other L-SBM diets (0.65, 0.75, 0.85%) exceeded the minimum SID ratio of tryptophan to lysine (0.18).
Classic health – amino acid nutrition principle
Our research confirms that the rate and efficiency of growth is governed by health status; healthier pigs grow faster and more efficiently than unhealthy pigs. It confirmed that the lysine (amino acids) requirement is higher for healthier than for unhealthy pigs. In addition, increasing lysine intake beyond the requirement for unhealthy pigs does not over-ride disease impaired-growth (Figures 1, 2). These landmark principles were established by Noel Williams and his Ph D advisor, Dr. Tim Stahly (Williams et al., 1997).
Our study extended concepts beyond their classic work, but in line with their hypothesis that FM in a product such as SBM may play a role in disease mitigation (personal communication, Dr. Stahly, 1998). The seemingly ‘inopportune’ infection by the SRD complex (co-infection with multiple respiratory pathogens) that we encountered created high-immune stressed pigs with profoundly impaired-growth. The observation that SBM (FM abundant source) had mitigating effects to counter the SRD complex is original and unusual; unusual in that the magnitude of response tends to be extraordinary for nutrition research.
Since SBM proved to have mitigating effects against SRD, it has dual and complementary functions as an ingredient: nutritional and tactical.
SBM improved nursery pigs’ ability to thrive
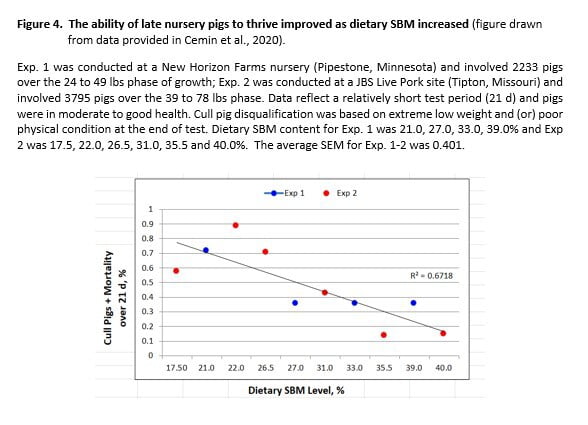
There are other examples of SBM improving the health status of pigs reared under commercial conditions (e.g., improved viability, reduced medical treatment). The report by Cemin and co-workers (2020) is especially noteworthy. Their collaboration with 2 production companies (JBS Pork, New Horizon Farms) afforded them access to 2 commercial farms and more than 6000 nursery pigs. Their primary objective was to observe the FCE response to increasing dietary SBM content (corn-SBM diet phase). Despite the narrow time frame of the test (21 d) and with pigs being in relatively good health, SBM reduced mortality and cull pigs (disqualified, extreme low weight or condition) in a dose-dependent manner (Figure 4). In other words, the ability of pigs to thrive in late nursery and early grower periods was positively related to SBM level.
Mechanisms for disease mitigating effects of SBM
A discussion of mode of action for the disease-mitigating effects of SBM is beyond the scope of this paper, but is the topic for our next article in this series. A few examples are provided here to support our hypothesis that systemic health was improved by dietary SBM during an infection with SRD complex.
We are indebted to human medical research for our understanding of disease mitigating mechanisms of soybeans. A variety of disease-uncoupling actions are exerted simultaneously by an equally diverse set of FM. Some mechanisms improve or balance the animal’s survival response to infection (no. 2, 3, 4 below), while other mechanisms operate to reduce pathogen concentration (no. 1). Disease mitigation mechanisms include (Chatterjee et al., 2018):
1. Anti-viral
Reduce virus replication and (or) increase viral clearance
2. Oxidation stress reduction
System responds to infection with free-radical production to combat many pathogens. Over-production of free radicals must be neutralized to prevent harm to animal cells and cell components.
3. Anti-inflammatory
Certain FM operate to control the level and balance of pro-inflammatory cytokines produced by the animals’ in response to infection. Over-production can be harmful to the animal.
4. Improve function of existing immune cells
5. Preserve intestinal barrier integrity
Lunasin, a polypeptide derived from SBM protein, exerts gut protective effects to protect barrier function.
Control of systemic disease requires multiple countering mechanisms that act in concert (e.g. anti-inflammatory, anti-oxidant) to reduce the damaging effects of pathogens. An effective counter to disease damage results in a durable or resilient state for normal function. The respiratory disease complex that we encountered in the Hanor study involved at least 3 known pathogens (virus, bacteria). Multiple pathogen infection tends to be the norm in commercial systems.
SBM – tactical option for SRD mitigation: DVM perspective
SBM is a tactical ingredient option in mitigating the severe growth and FCE impairing effects of SRD. We applied the term prescriptive to this ingredient as a cuing device for veterinary and nutrition decision-makers to be mindful of the mitigating benefits of SBM during SRD, in addition to its’ amino acid and energy value. Replacing SBM with alternative protein sources can be made an equal trade for nutrition, but this is not true for FM even among other legumes. SRD imposes significant hidden costs in FCE loss, increased medical treatment and potentially fewer pigs that thrive in late nursery through early growth. These costs are hidden because they are ‘folded’ into barn ‘closeout’ data, but they can add $3-6/pig to cost (range $2-9).
SRD complex is prevalent in many North American systems during January – April, when barns are closed for cold weather (see endnote for respiratory disease pathogen). Influenza is also a problem in the fall for some systems, but the economic cost of winter - spring SRD tends to be more persistent.
The financial value of tactical SBM use can be determined by comparing performance for (1) system diets and (2) prescriptive diets (system diets plus 30% more SBM in each diet to finish). Diets need to be equally applied by site and sex, using large numbers and in the time frame specified by the Veterinarian(s). Key measures are tracked to compute the financial outcome (e.g., 1000 pig basis), including full-value (FVP) and cull-value pig market revenue, total diet cost and medication cost.
Carcass weight is a priority measure. Although growth rate is reflected in FVP carcass pounds, the diet effect on population carcass weight is important because a decline can begin to emerge even prior to summer heat stress. The motive is to see if the system is experiencing a leading edge to summer carcass weight dip (later article in this series). Carcass weight decline can begin to appear in April when heat is not yet a problem and becomes worse during summer. The reason for a premature decline in carcass weight can be confusing, but it could be due to unchecked SRD impaired-growth. Summer carcass weight dip is a very costly problem; one that is preventable provided that pre-summer carcass weight decline is controlled.
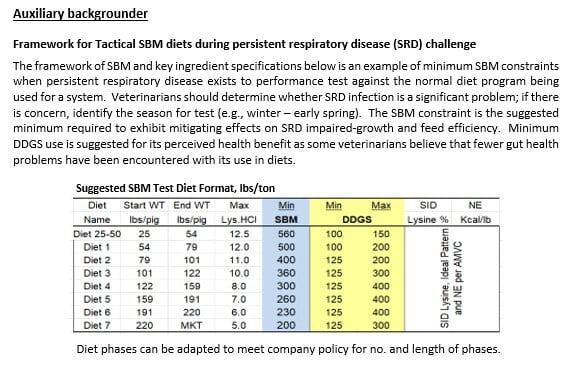
An example of ingredient constraints (SBM, DDGS) for a field test of standard and prescriptive or tactical SBM diets is shown in the Auxiliary section below (provided courtesy of The Hanor Company).
Key Conclusions
Respiratory disease (SRD) impairs feed conversion and daily gain by at least 8 and 10%
Increasing dietary SBM above typical levels mitigates FCE and ADG loss during SRD
SBM is both a nutritional and prescriptive ingredient
Cost-effective periods for elevated SBM may be during Winter – Spring seasons, when barns are closed tight and when SRD is often a problem
Endnote
The most problematic swine respiratory diseases include Mycoplasma hyopneumonia, Porcine reproductive and respiratory syndrome (PRRSv), Influenza A virus in swine, Streptococcus suis, Hemophilus parasuis, and Actinobacillus suis pleuropneumonia.
References
Sharma, R., S. Kumar, V. Kumar and A. Thakur. 2019. Comprehensive review on nutraceutical significance of phytochemicals as functional food ingredients for human health management. J. Pharma. Phytochem. 8 (5):385-395. http://doi.org/10.22271/phyto.2019.v8.i5h.9589
Chatterjee, C., S. Gleddie and C-W Xiao. 2018. Soybean bioactive peptides and their functional properties: Review. Nutrients 10 (9):1211-1227. http://dx.doi.org/1.3390/nu10091211
Smith, B.N. and R.N. Dilger. 2018. Immunomodulatory potential of dietary soybean-derived isoflavones and saponins in pigs. J. Anim. Sci. 96:1288-1304. http://doi.org/10.1093/jas/skyy036
Johnston, M.E., R.D. Boyd, C. Zier-Rush, and C.E. Fralick. 2010. Soybean meal level modifies the impact of high immune stress on growth and feed efficiency in pigs. J Anim. Sci. 88 (E-Suppl. 3):174.
Boyd, R.D., M.E. Johnston and C. Zier-Rush. 2010. Soybean meal level modulates the adverse effect of high immune stress on growth and feed efficiency in growing pigs. Proc. 71st Minn. Nutrition Conf. pp. 167-174. U. Minn., St. Paul MN. https://hdl.handle.net/11299/204236x
Cemin, H. S., H. E. Williams, M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. Derouchey, K. F. Coble, B. A. Carrender, and M. J. Gerhart. 2020. Estimate of the energy value of soybean meal relative to corn based on growth performance of nursery pigs. J. Anim. Sci. Biotech 11:70-78. https://doi.org/10.1186/s40104-020-00474-x
National Research Council (NRC). 2012. Nutrient requirements of swine 11th revised edition. National Academy Press, Washington, DC, USA. https://doi.org/10.17226/13298
Boyd, R.D. and A.M. Gaines. 2023. Soybean meal NE value for growing pigs is greater in commercial environments. Feedstuffs, August digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-august-2023/page/2/1
Parra, M.D., P. Fuentes, F. Tecles, S. Martinez-Subiela, J.S. Martinez, A. Munoz and J.J. Ceron. 2006. Porcine acute phase protein concentrations in different diseases in field conditions. J. Vet. Med. B 53:488-493.
Johnson, R.W. 1997. Inhibition of growth by pro-inflammatory cytokines: An integrated view. J. Anim. Sci. 75:1244-1255
Spurlock, M.E. 1997. Regulation of metabolism and growth during immune challenge: An overview of cytokine function. J. Anim. Sci. 75:1773-1783.
Williams, N.H., T.S. Stahly and D.R. Zimmerman. 1997. Effect of chronic immune system activation on the rate, efficiency, and composition of growth and lysine needs of pigs fed from 6 to 27 kg. J. Anim. Sci. 75:2463-2471. https://doi.org/10.2527/1997.7592463
R. Dean Boyd, Ph D. is an adjunct professor of Animal Nutrition at North Carolina State U. and Iowa State U.; Michael E. Johnston, MS is an animal nutritionist and research scientist, United Animal Health; James L. Usry, Ph D. is research scientist and principle managing partner of Ani-Tek Group LLC; Paul Yeske, DVM MS is the senior member of the Swine Vet. Center team, St. Peter, Minnesota; Aaron M. Gaines, Ph D. is an animal nutritionist and co-founder and managing partner of Ani-Tek Group LLC.