Functional compounds in soybean meal: Implications for pig health and physiology
This article is the 4th in a series highlighting the unique value of soybean meal to swine nutrition and health.
By Amy L. Petry, Brooke Bowen, Lisa Weaver, and R. Dean Boyd
Introduction
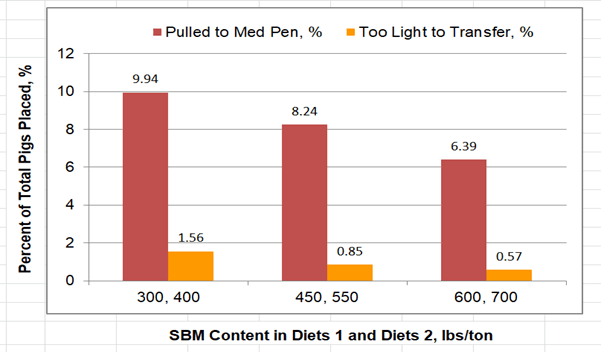
Previous papers in this series (1, 2) show apparent extra-nutrient effects of soybean meal (SBM) on pig performance in the commercial environment. SBM’s benefits have been documented for growth, viability, and medical intervention of pigs growing under conditions of both high and low immune stress. For example, dietary SBM contributed to the mitigation of severe growth-impairing effects of respiratory disease in finishing pigs. In a subsequent study, the need for medical intervention declined and the ability to thrive (live, grow) improved in Porcine Reproductive and Respiratory Syndrome virus (PRRSv)-challenged nursery pigs in response to increasing dietary SBM content (3), as illustrated in Figure 1. Production improvements with low immune stress were reported by Cemin and coworkers (4) who reported a linear improvement in the ability of nursery pigs to thrive as dietary SBM level increased (6000 pigs, 2 systems).
A noteworthy phenomenon emerged during validation of SBM net energy (NE) in commercial settings. SBM NE values exceeded a recent calorimetric estimate for SBM by at least 20% (2). This result was unexpected but consistent across 5 commercial studies that was conducted by 3 research groups on 2 continents; each conducted under conditions of subclinical disease stress. These results do not necessarily conflict with classically-derived NE estimates, but may represent conservation of maintenance energy that otherwise might have been used to counter pathogens or other stressors in the commercial environment.
* Amy L. Petry, Ph.D. is an Assistant Professor of Monogastric Nutrition in Animal Science, University of Missouri, Columbia; Brooke Bowen has an M.S in Animal Science from Texas Tech University and studied the mechanism of functional compounds in soybeans under Dr. Petry; Lisa Weaver, Ph.D. in Plant Biochemistry, is Research Director with Smithbucklin; R. Dean Boyd, Ph.D., is adjunct professor of Animal Nutrition at North Carolina State University. and Iowa State University.
While SBM is well known as a quality source of protein, approximately 40% of processed SBM is something other than protein, moisture and oil (5). It has been hypothesized that this fraction contains extra-nutrient components that may be involved in mitigating respiratory disease effects and improving the energy partitioning for growth in commercially-reared pigs (1, 2). SBM is an abundant and diverse source of functional compounds (6) in the non-protein fraction, as well as bioactive peptides that arise from parent proteins (6). Peptides that are encrypted in seed proteins arise by digestion and these protein fragments are not active until released from the protein (7).
This paper provides a targeted review of the functional compounds in SBM that may be involved in its disease mitigating effects in swine, and potentially the improvement of energy partitioning for growth under conditions of low immune stress (or subclinical disease).
Emerging evidence for value of SBM functional compounds in pigsMechanistic research on soybean functional compounds in food animals is in its infancy. However, important findings are beginning to emerge, showing how functional compounds in SBM could be acting therapeutically in the commercial sector to reduce impaired-growth, reduce medication needs and improve viability under systemic disease challenge.
Dilger Lab – University of Illinois
A number of studies have been published by the Ryan Dilger lab using young pigs infected with PRRSv to create the health disturbances commonly encountered in commercial practice (impaired growth, mortality). They investigated the rate of recovery from infection with and without isoflavones (ISF) in their diet. The level and proportion of ISF added was similar to a diet having 20% SBM. Isoflavones have been shown to have anti-inflammatory, antiviral and anti-oxidative effects largely within the GI tract (8).
PRRSv-infected, weaned pigs that received isolated dietary ISF (more bioavailable than in SBM) exhibited an improved adaptive immune response compared to those not receiving ISF (9). This promising result was followed by a long-term study (161 d) to determine whether feeding PRRSv-infected pigs diets with ISF isolates would benefit growth and (or) viability compared to counterparts fed diets without ISF addition. The study began 7 d post-weaning and ended at approximately 125 kg (9).
The PRRSv infection was complicated by a secondary bacterial infection (Streptococcus suis), which is consistent with commercial experience. During the period of active PRRSv infection (inoculation to 34 d post-infection, PI), growth rate was equally impaired (-39.6%) for both PRRSv-treatments compared to non-infected control pigs. The same was true for the gain to feed ratio (-21.0% FCE). Growth rate and FCE did not differ during the period of recovery (34 to 90 d PI) or after viral clearance (90 to 161 d PI).
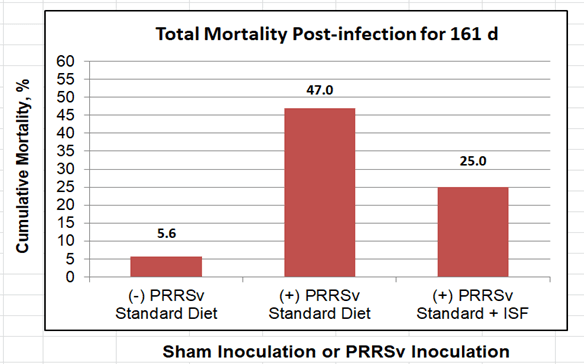
Although dietary ISF did not affect either growth rate or FCE, it did improve the acute immune response, which was followed by reduced time for PRRSv clearance from the system (102 d vs 123 d PI). Dietary ISF also elicited more robust anti-PRRSv neutralizing antibody production earlier in the disease progression. The more rapid clearance of PRRSv by pigs receiving ISF may partially contribute to mechanisms leading to fewer infection-related deaths (25% mortality) over the entire growth period, compared with infected pigs not receiving ISF (47% mortality). Pigs in the latter group also had more severe secondary bacterial infections. The pattern of mortality for the 161 d period is shown in Figure 2.
These studies significantly advanced our understanding of soybean ISF value in countering the ubiquitous respiratory pathogen PRRSv. Feeding ISF to PRRSv -infected pigs led to an earlier adaptive immune response to neutralize PRRSv, which resulted in more rapid clearance and less severe secondary bacterial infection. Once ISF-fed pigs entered the PRRSv recovery phase, they did so more completely.
The net result of ISF promoting greater immune protection to the primary and secondary pathogens was to reduce overall mortality by one-half (25%) compared to infected pigs not receiving ISF (47%). This is a high value production outcome.
Petry Lab – University of Missouri
Within Petry’s Lab, researchers evaluated the effect of isolated SBM functional compounds in nursery pig diets, relative to intact SBM (10), in order to clarify the role of these compounds on nursery pig gastrointestinal physiology, systemic health, and inflammatory markers and aid in elucidating their mechanism of action in pigs. The diets included a high SBM (25%+) diet as a positive control; a negative control without SBM but supplemented with soy protein concentrate and isolate; and four other diets fortified with different soy compounds including isoflavones, soybean phospholipids, soluble fiber, oligosaccharides, and a soy peptide known as lunasin in a concentration similar to that provided by the SBM control. The results revealed a 30% decrease in white blood cells, particularly neutrophils, in pigs fed diets with isolated isoflavones and the SBM control, indicating an immune cell modulatory effect. Further cell-based work suggested that these dietary components might dampen the body's inflammatory response through a mechanism involving isoflavones’ impact on antioxidative enzymes and decreasing the production of reactive oxygen species by leukocytes (10).
Another interesting finding was an improvement in systemic and gastrointestinal antioxidant capacity with diets including soy lecithin and phytosterols. This was coupled with a decrease in malondialdehyde (MDA), a byproduct of lipid peroxidation, which is commonly associated with cellular damage and stress. These changes suggest that soy components may bolster antioxidant defense mechanisms, thus protecting tissue integrity and function. Taken together, the study's insights suggest that soy's functional components, particularly its lipids and polyphenol components, could confer health benefits in young pigs. The effects observed in pigs may parallel the cardiovascular and immune system benefits that these compounds have in humans (6), pointing to a similar therapeutic potential in animal diets.
The remainder of this article is a brief, targeted review of the key functional compounds in SBM and their potential mechanisms of action in swine (Figure 3). Much of this review relies on the human nutrition and medicine research for an understanding of their potential physiological effects.
Concept of Functional Foods and Compounds
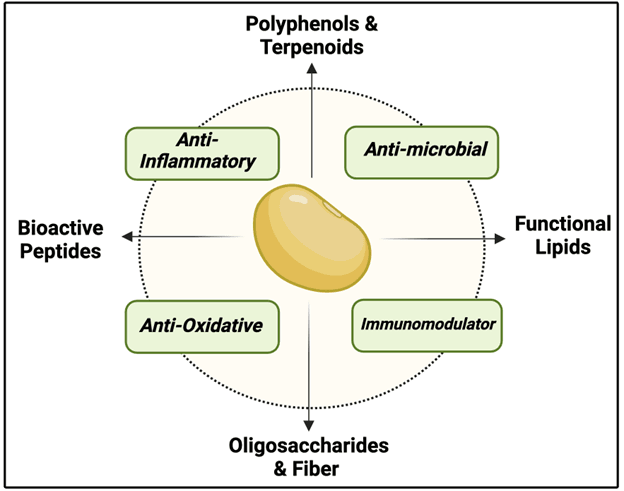
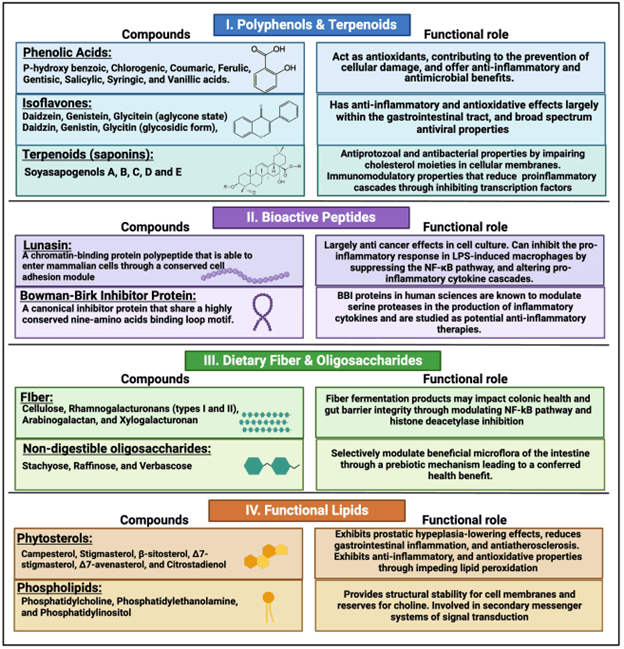
“Functional foods” have emerged as a critical concept in human nutrition, with a growing body of clinical evidence supporting their role in preventing and managing chronic illnesses. Soybeans have been recognized as a functional food due to their dense concentration of functional compounds (11). A functional or phytochemical compound is a naturally derived, dietary compound that is regularly consumed and impacts a particular physiological process (12). Functional compounds are not considered nutrients, as their lack does not correlate with any specific deficiency. Many cell culture and population studies within the human sciences have identified soybeans as a source of at least 14 functional compounds (6), including phytic acid, peptides, triterpenes, phenolics and flavonoids, fatty acids, carotenoids, coumarins, protease inhibitors, oligosaccharides, and dietary fibers. These compounds can act alone or synergistically with the gut microbiome to create metabolites, which impact health and physiology. While certain compounds, such as protease inhibitors and lecithin, can contribute positively to human health by aiding in weight management and reducing atherosclerosis, they can hinder nutrient uptake and growth performance in swine depending on their dietary concentration (5). However, processed SBM contains four critical categories of functional compounds that may contribute to health-oriented pork production objectives, specifically polyphenols and terpenoids, bioactive peptides, dietary fiber and oligosaccharides, and functional lipids (Figure 4).
Functional Compounds in SBM: Classification and Description
These four categories represent approximately 70% of the functional compounds found in soybeans that have recognized effects in human nutrition, and are most prevalent in soybean meal.
1. Polyphenols and terpenoids including phenolic acids, isoflavones, saponins
2. Bioactive peptides, usually generated by designed, incomplete digestion
3. Dietary fiber and oligosaccharide
4. Functional lipids, including phytosterols and phospholipids
1. Polyphenols and terpenoids
Soybeans produce polyphenolic compounds, through their secondary metabolism, which include phenolic acids, isoflavones, and saponins. These are the most abundant and diverse functional compounds in soybeans. Polyphenols within the bean are multifunctional compounds, essential for defense against plant pathogens while also contributing to the plant's overall health and environmental adaptation (13).
Phenolic Acids: Eight phenolic acids (p-hydroxy benzoic, chlorogenic, coumaric, ferulic, gentisic, salicylic, syringic, and vanillic acids) are consistently present in soybeans (6). These compounds can be found in an accessible form, a free conjugate form, or as a bound component, and are all derived from phenylalanine (14). SBM is 37 to 58% more concentrated in phenolic acids than whole soybeans and has a 60% greater antioxidant capacity (15, 16). This is because protein is concentrated in SBM production, and most phenolic acids are part of the protein unit of the bean. However, the bioavailability of bound phenolic acids, which must be released into the gastrointestinal tract to be biologically active, still needs to be established in swine. In vitro, monogastric digestion models suggest an 8 to 28% bioavailability of p-hydroxy benzoic, chlorogenic, and p-coumaric acids from soybeans (17). Several phenolic acids also serve as intermediate metabolites in the phenylpropanoid pathway to produce flavonoids such as isoflavones.
Isoflavones: Isoflavones, classified as flavonoids, have a structural resemblance to 17β-estradiol and include three molecules—daidzein, genistein, and glycitein—each of which presents in four structural motifs in soybeans. They are bioavailable in their aglycone state (called daidzein, genistein, glycitein). Conversely, their glycosidic form (daidzin, genistin, glycitin) is bound to glucose and is not bioavailable (6). These glycosides can acquire acetyl or malonyl groups, forming acetylglucosides or malonylglucosides, respectively. Isoflavones bound to a carbohydrate motif can be hydrolyzed by β-glycosidases, which are produced by the host or microbiota. Once hydrolyzed, they are bioavailable and can be absorbed predominately in the small intestine or metabolized further by microbiota. Most absorbed isoflavones are metabolized by enterocytes, with only a fraction entering circulation (<5%) (18). Genistein and daidzein, in their free (aglycone) and glycosidic (glycone) forms, represent 60% and 30% of the total isoflavone content of soybean, with glycitein present at <10%. Isoflavones contribute to plant-microbial interactions by mitigating stress-induced reactive oxygen species (ROS) and facilitating defense against plant pathogens (13). Because of their function in plants, isoflavone concentration and composition is known to vary considerably across cultivars, locations, and growing conditions.
Saponins: Saponins are surfactant terpenoid compounds consisting of a sapogenin core attached to one or more carbohydrate moieties and found predominately in the germ. Five distinct sapogenins (A-E) have been characterized in soybeans, each potentially glycosylated with three monosaccharides such as galactose, arabinose, rhamnose, glucose, xylose, or glucuronic acid (19). Due to their complex interactions with carbohydrates, they are challenging to analyze and isolate. Like isoflavones, their bioavailability depends on then being hydrolyzed from the glycone to aglycone form. Uniquely, a self-micellization process increases their solubility, enhancing their absorption by enterocytes (20). Soybeans rely on saponins mainly for defense, as these compounds provide antimicrobial benefits against fungi and bacteria for the plant. Like isoflavones, the concentration of saponins depends on genetic source, cultivation conditions, and processing methods.
2. Bioactive Peptides
Soybean meal protein is a composite of protein types or families, 80-90% of which are glycinin and beta-conglycinin (21). The mixture of protein types in SBM varies slightly with genetics and with the environment where the soybeans are grown (22). These two major proteins undergo enzyme cleavage during digestion (by endo- and exo-peptidases) and through gut microbiome activity after ingestion. Protein cleavage can also occur if soybean meal is fermented before being added to the diet. Since proteins are chains of amino acids, when they are cleaved the result is individual amino acids and a mixture of smaller peptides. Those peptides with amino acid to amino acid linkages for which an endopeptidase does not exist appear to represent a designed outcome (23).
The fact that some peptides are bioactive themselves, and have effects beyond just delivering amino acids, has been known for some time. Bioactive peptides are usually not active until released from the parent protein, and are generally between two and twenty amino acids long (24). Lunasin and the Bowman-Birk inhibitors (BBi) are small proteins which are self-existent bioactive peptides (oligo-peptides), larger than other bioactive peptides identified from soybean (24). Lunasin has long been known for anti-cancer activity and has anti-inflammatory activity (25), while BBi has been associated with antiviral and anti-inflammatory activity (26). With the exception of lunasin and BBi, the activity of proteases is required to release the active peptide, so the ability to consistently produce the same mixtures of peptides through the digestive process will likely depend on species, development stage, and gut microbiome composition (28, 29, 30)
3. Dietary Fiber and Oligosaccharides
Carbohydrates constitute approximately 30-35% of soybean’s dry matter content (31). These carbohydrates are predominantly nonstructural, including a minor fraction of starch, sugars, oligosaccharides, and dietary fiber. Solvent-extracted SBM is particularly noted for its dietary fiber and prebiotic oligosaccharides content, averaging 12±6% total dietary fiber and 5±2% non-digestible oligosaccharides (32). The fermentable fiber in SBM is characterized by a variety of sugars such as galactose, arabinose, and fructose, as well as uronic acids like galacturonic acid, with notable quantities of non-digestible galactooligosaccharides (NDOs) including predominantly stachyose, with smaller amounts of raffinose, and verbascose (31, 33). While dietary fiber can impede nutrient digestibility, its fermentable components are valued for their beneficial effects on gastrointestinal health (34). Nevertheless, NDOs may exert antinutritive effects in young pigs, potentially causing gastrointestinal disturbances and resulting in loose fecal consistency (5).
4. Functional Lipids
The functional lipid content of SBM, such as phytosterols and phospholipids, is subject to variation based on processing conditions and growing intensity (35). These lipids are present in lower quantities and exhibit considerably more variability than other functional compounds in SBM, conceivably playing a lesser role in its functional attributes. However, soy lecithin, a phospholipid, can be reintroduced into SBM through 'addbacks' during the soy oil refining process, resulting in fluctuating concentrations in end products. Phospholipids, integral to cell membrane composition, include two fatty acids, a phosphate group, and a glycerol backbone. In soybeans, prevalent phospholipids include phosphatidylcholine, phosphatidylethanolamine, and phosphatidylinositol (36). Additionally, phytosterols, which are plant sterols analogous in structure to cholesterol, are present in soybeans as six isomers (campesterol, stigmasterol, β-sitosterol, ∆7-stigmasterol, ∆7-avenasterol, and citrostadienol) comprising approximately 0.4% of the total weight (37). While most soy phytosterols are extracted alongside the oil, soybean products that have undergone expansion or extrusion and thus have a higher ether extract content are considered potential sources of these functional compounds.
Functional Compounds: Potential Mechanisms of Action
Biological Functions of SBM Functional Compounds of Importance to Food Animals
In the human sciences, there are a plethora of papers on the functional mechanisms of soy-based foods, including reviews by Messina et al. (11) and Basson et al. (6), as well as a compendium of research featured in a special edition of Frontiers in Nutrition (38). This section aims to synthesize the proposed mechanisms of soy's functional compounds as inferred from various domains, including cell culture experiments and disease challenge models. We recognize the challenges in translating these mechanisms to swine in a commercial context. While not ideal, these insights establish a foundational knowledge base to further investigate and understand the modulatory effects of SBM observed in commercial conditions (Figure 5).
1. Functional aspects of polyphenols, flavonoids and terpenoids
The antioxidant, antimicrobial, and anti-inflammatory properties of polyphenols, flavonoids, and terpenoids have garnered significant attention in both human and animal science research. Microbial metabolism can transform the more complex polyphenol structures into potent phenolic metabolites, such as equol and salicylic acid (39). However, the bioavailability of polyphenols and terpenoids in pigs remains poorly understood. These compounds must first be hydrolyzed from their plant matrix, through processing or digestion, and then absorbed in the gut. Research in humans and mice suggests that the large intestine is the primary site of absorption for polyphenols and isoflavones, and the observed reduced plasma concentrations post-consumption indicate that their effects are localized to intestinal cells (40). Thus, the functional roles of soy polyphenols are likely localized to the gastrointestinal tract, and further research on bioavailability is warranted. The following are key potential physiological mechanisms attributed to bioavailable soy polyphenols and terpenoids.
Soy phenolic and flavonoid compounds act as antioxidants through electron donation and metal chelation, a process influenced by the special arrangement of their functional groups, protecting against oxidative damage to proteins, nucleic acids, and lipids (41). Research has highlighted the effectiveness of antioxidant properties of phenolic acids and isoflavones in both cellular and enzymatic systems (42, 43). Specifically, isoflavones combat oxidation by neutralizing free radicals and inhibiting singlet oxygen without producing additional radicals (44). This unique property allows it to recycle as an antioxidant more effectively than many other functional compounds. Dietary soy isoflavones in nursery pigs have improved jejunal antioxidant levels and regulated gut barrier integrity, potentially through a means of mitigating ROS (45, 10). Additionally, equol, a microbial metabolite produced from daidzein, has greater antioxidant activity than its parent compound and is more readily absorbed into the bloodstream (46).
Soy isoflavones possess complementary anti-inflammatory and antioxidant properties. Their primary mechanism involves immunomodulatory actions through inhibiting tyrosine-specific protein kinases and NF-κβ transcription, which are key players in regulating inflammation and producers of cellular oxidants (47, 7). Saponins can also mediate NFκB transcription factors that mitigate proinflammatory responses (48). Likewise, phenolic compounds' anti-inflammatory properties suppress inflammatory mediators like prostaglandins and nitric oxide, potentially reducing inflammation (49). Diet supplementation with phenolic-enriched soy isoflavones has been linked to enhanced immune function, marked by reduced diarrhea incidence and lower endotoxin levels in lipopolysaccharide (LPS)-induced, immune-challenged nursery pigs (50). They also activate mononuclear cells, bolstering the phagocytic response through MAPK and NF-κB pathway modulation in mice models (51). Furthermore, cell culture work suggest phenolic acids may contribute to gut health by inhibiting the COX-2 enzyme, implicated in inflammatory responses, and thus, decreasing pro-inflammatory prostaglandins in activated cells (43).
Isoflavones have demonstrated antiviral properties across various viruses, by interfering with different stages of the virus life cycle including viral entry, replication, protein synthesis, and virus envelope formation (52). Genistein is particularly noted for its broad-spectrum antiviral efficacy against various types of viruses, including those with differing envelope structures and genetic materials (53). The effectiveness of flavonoids and isoflavones in antiviral defense, however, depends on the dosage used, ranging from physiological to supraphysiological levels.
An in-depth discussion of the potential antiviral and antibacterial mechanisms of soy isoflavones and saponins, a subclass of terpenoids, is found in the comprehensive analysis by Smith and Dilger (8). Their findings suggest that administering soy-derived isoflavones or saponins to infected animals could potentially enhance recovery, lower mortality rates, and stabilize performance in
specific conditions.
2. Functional aspects of bioactive peptides
Bioactive peptides may have both local and systemic effects. With respect to the latter, dipeptides (two amino acids) are easily absorbed, but there is evidence that longer peptides are taken up by various mechanisms (54).
Soy bioactive peptides target localized cell proliferation, so may be anticarcinogenic (55).
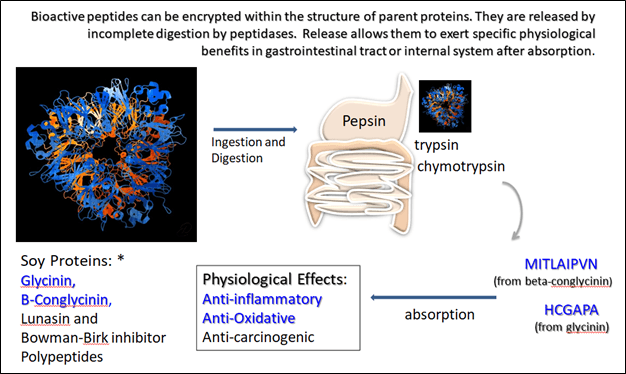
The generation of bioactive peptides from major proteins is shown in Figure 6. In this example, a peptide is released through digestion from the beta-conglycinin (MITLAIPVN) and glycinin (HCGAPA) protein families. The first peptide stimulates immune cell function, and the latter stimulates phagocytosis by macrophages and leukycytes (24). In total, the collection of peptides from soy protein improves immune cell function, reduces oxidative stress, and is anti-inflammatory (e.g. block pro-inflammatory cytokines) (24). This process of unveiling bioactive amino acid chains from a larger protein is not limited to soybean; it is also characteristic of other proteins such as whey (56).
In addition to the cleavage products of larger proteins, small peptides found in soybean can also be a source of bioactive peptides. Soy lunasin has long been noted for its anti-cancer and anti-inflammatory activity. The anti-inflammatory activity may be due to a tripeptide, arginine-glycine-aspartate, found within lunasin, which could be released by digestion and then disrupts the inflammatory cascade (25). Finally, the soybean Bowman-Birk trypsin inhibitor (BBi) is often considered a bioactive peptide. This small protein, almost twice as large as lunasin, has been shown to have anti-inflammatory and anti-viral activities (26, 27). BBi is believed to be quite stable, due to its multiple di-sulfide bonds.
Unlike other bioactive peptides which are released by digestion, BBi’s ability to inhibit proteases may be responsible for its activity. The inflammatory response involves serine proteases, and inhibition of these proteases results in reduced proinflammatory cytokine release (26). Likewise, the inhibition of key enzymes involved in virus replication may be the source of BBi’s antiviral activity (26).
3. Functional aspects of non-digestible oligosaccharides (NDOs) and dietary fiber
Pigs are unable to digest soy fiber or NDOs, and instead depend on microbial fermentation. These NDOs can exhibit a prebiotic effect in the gastrointestinal tract at low concentrations. The International Scientific Association for Probiotics and Prebiotics (57) defines prebiotics as “substrates selectively utilized by host microorganisms that confer a health benefit”. It is crucial to recognize that the impact of NDOs on health and productivity varies with their dietary concentration. Above 3%, NDOs may negatively influence the digestibility and energy value of SBM; however, within the concentrations typical in corn-SBM-based grower diets, they may serve as prebiotics, promoting the proliferation of beneficial bacteria like Lactobacillus and Bifidobacterium (31,58). Moreover, soybean NDOs may inhibit the production of putrefactive compounds from protein fermentation in the large intestine (59). This modulation of gut microbial communities by soybean NDOs may leverage a prebiotic-like effect to reduce protein-derived catabolites in swine's intestinal luminal content (60). Additionally, the fermentation of galactooligosaccharides is known to stimulate Bifidobacteria growth in humans, thereby altering colonic microbiota (61). Bifidobacteria are known for improving gut barrier integrity, vitamin synthesis, and immune function enhancement (62).
4. Functional aspects of phytosterols and phospholipids
In humans, phytosterols and phospholipids are valued for their contribution to cell membrane integrity, heart and liver health, and antioxidant effects (64). However, when consumed by young animals and fish, they can be antinutritive (63). Soybean phospholipids, including glycerophospholipids and phosphatidylcholine, can aid in forming cell membranes and the intestinal mucus layer. Phospholipids play a crucial role in the composition of intestinal mucus, existing as a continuous layer on the luminal side of the mucus gel. They are present within the mucus in the form of liposome-like aggregates, and form a monolayer on the surface of mucosal cells. Clinical applications of phosphatidylcholine in humans have shown anti-inflammatory properties, largely driven by modifying the prostaglandin pathways and inhibiting TNF-α-induced inflammatory responses (65). The phytosterols found in soybeans, such as β-sitosterol and campesterol, are beneficial for lowering cholesterol and may also influence immune responses related to colitis (66). Human studies indicate that phytosterols may have anti-inflammatory and antioxidant properties, potentially reducing the inflammatory response of immune cells (67). Currently, there is a dearth of knowledge on the direct effects of phytosterols and phospholipids found in SBM, as compared to contained in soy oil.
Conclusion
The collective body of research presented in this paper highlights the multifaceted role of compounds in soybeans, as recognized by other sectors. While SBM is known in swine nutrition for its protein content, it is the non-protein fraction—rich in polyphenols, terpenoids, bioactive peptides, fiber, functional lipids and other functional compounds—may play a role in modulating health outcomes in pigs and may be considered as complementary nutrition factors. Data indicate possible benefits of SBM in disease resilience and physiological responses, yet the mechanisms of action remain to be fully elucidated, and bioavailability will be a key factor in efficacy.
References
Boyd, R.D., M.E. Johnston, J. Usry, P. Yeske and A. Gaines. 2023. Soybean meal mitigates respiratory disease-impaired growth in pigs. Feedstuffs, October digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-october-2023/page/2/1.
Boyd, R.D. and A.M. Gaines. 2023. Soybean meal NE value for growing pigs is greater in commercial environments. Feedstuffs, August digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-august-2023/page/2/1
Zier-Rush, C., S. Smith, J. Picou, E. van Heugten and R.D. Boyd. 2015. High SBM level reduces medical treatment of PRRS affected weaned pigs fed early nursery diets. Hanor Tech. Memo H-2015-07. https://doi.org/10.13140/RG.2.2.35498.39366
Cemin, H. S., H. E. Williams, M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. Derouchey, K. F. Coble, B. A. Carrender, and M. J. Gerhart. 2020. Estimate of the energy value of soybean meal relative to corn based on growth performance of nursery pigs. J. Anim. Sci. Biotech 11:70-78. https://doi.org/10.1186/s40104-020-00474-x
Choct, M., Y. Dersjant-Li, J. Mcleish and M. Peisken, 2010. Soy oligosaccharides and soluble non-starch polysaccharides: A review of digestion, nutritive and antinutritive effects in pigs and poultry. Asian-Aust. J. Anim. Sci. Vol. 23. No. 10:1386- 1398
Basson AR, Ahmed S, Almutairi R, Seo B, Cominelli F. Regulation of Intestinal Inflammation by Soybean and Soy-Derived Compounds. Foods. 2021; 10(4):774. https://doi.org/10.3390/foods10040774
Sanchez, A. and A. Vazquez. 2017. Bioactive peptides: A review. Food Qual. Safety 1:29-46. Doi:10.1093/fqs/fyx00
Smith, B.N. and Dilger, R.N., 2018. Immunomodulatory potential of dietary soybean-derived isoflavones and saponins in pigs. Journal of Animal Science, 96(4), pp.1288-1304.
Smith, N.S., M.L. Oelschlager, M.S.A. Rasheed, R.N. Dilger. 2020. Dietary soy isoflavones reduce pathogen-related mortality in growing pigs under porcine reproductive and respiratory syndrome viral challenge. Journal of Animal Science, 98(4), February 2020. https://doi.org/10.1093/jas/skaa024.
Bowen, B., 2023. Evaluating the relationship of bioactive functional compounds innate to soybean meal on nursery pig performance, gut physiology, nutrient digestibility, and systemic health. Thesis. Texas Tech University.
Messina, M., Duncan, A., Messina, V., Lynch, H., Kiel, J., & Erdman Jr, J. W. (2022). The health effects of soy: A reference guide for health professionals. Frontiers in Nutrition, 1837.
Dillard, C.J. and German, J.B., 2000. Phytochemicals: nutraceuticals and human health. Journal of the Science of Food and Agriculture, 80(12), pp.1744-1756.
Singh, S., Kaur, I. and Kariyat, R., 2021. The multifunctional roles of polyphenols in plant-herbivore interactions. International Journal of Molecular Sciences, 22(3), p.1442.
Xu, Minwei, Jiajia Rao, and Bingcan Chen. Phenolic Compounds in Germinated Cereal and Pulse Seeds: Classification, Transformation, and Metabolic Process. Critical Reviews in Food Science and Nutrition 60, no. 5 (March 8, 2020): 740–59. https://doi.org/10.1080/10408398.2018.1550051
de Oliveira Silva, F. and Perrone, D., 2015. Characterization and stability of bioactive compounds from soybean meal. LWT-Food Science and Technology, 63(2), pp.992-1000.
Xu, B. and Chang, S.K., 2008. Total phenolics, phenolic acids, isoflavones, and anthocyanins and antioxidant properties of yellow and black soybeans as affected by thermal processing. Journal of Agricultural and Food Chemistry, 56(16), pp.7165-7175.
Zhu, L., Li, W., Deng, Z., Li, H. and Zhang, B., 2020. The composition and antioxidant activity of bound phenolics in three legumes, and their metabolism and bioaccessibility of gastrointestinal tract. Foods, 9(12), p.1816.
Larkin, T., Price, W.E. and Astheimer, L., 2008. The key importance of soy isoflavone bioavailability to understanding health benefits. Critical Reviews in Food Science and Nutrition, 48(6), pp.538-552.
Zhao, G., Zhu, L., Yin, P., Liu, J., Pan, Y., Wang, S., Yang, L., Ma, T., Liu, H. and Liu, X., 2022. Mechanism of interactions between soyasaponins and soybean 7S/11S proteins. Food Chemistry, 368, p.130857.
de Hierro, J.N., Herrera, T., Fornari, T., Reglero, G. and Martin, D., 2018. The gastrointestinal behavior of saponins and its significance for their bioavailability and bioactivities. Journal of Functional Foods, 40, pp.484-497.
Natarjan et al. Characterization of Storage Proteins in Wild (Glycine soja) and Cultivated (Glycine max) Soybean Seeds Using Proteomic Analysis J. Agric. Food Chem. 2006, 54, no 8, 3114–3120 https://doi.org/10.1021/jf052954k
Assefa et al. Assessing Variation in US Soybean Seed Composition (Protein and Oil) Front Plant Sci. 2019, 10, https://www.frontiersin.org/articles/10.3389/fpls.2019.00298
Kitts, D.D. and Katie Weiler. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Curr Pharm Des. 2003;9(16):1309-23. doi: 10.2174/1381612033454883
Chatterjee, Cynthia, Stephen Gleddie, and Chao-Wu Xiao.. Soybean Bioactive Peptides and Their Functional Properties Nutrients 2018 10, no. 9: 1211. https://doi.org/10.3390/nu10091211
Lule, V.K., Garg, S., Pophaly, S.D., Hitesh, and Tomar, S.K. (2015), Potential Health Benefits of Lunasin: A Multifaceted Soy-Derived Bioactive Peptide. Journal of Food Science, 80: R485-R494. https://doi.org/10.1111/1750-3841.12786
Agata Gitlin-Domagalska , Aleksandra Maciejewska and Dawid Debowski, Bowman-Birk Inhibitors: Insights into a Family of Multifunctional Proteins and Peptides with Potential Therapeutical Applications (2020) Pharmaceuticals 13, 421; doi:10.3390/ph13120421
de Almeida Barros, B., da Silva, W.G., Moreira, M.A. et al. In silico characterization and expression analysis of the multigene family encoding the Bowman–Birk protease inhibitor in soybean. Mol Biol Rep 39, 327–334 (2012). https://doi.org/10.1007/s11033-011-0742-1
Chong Wang, Fan Zhao, Yun Bai, Chunbao Li, Xinglian Xu, Karsten Kristiansen, Guanghong Zhou, Effect of gastrointestinal alterations mimicking elderly conditions on in vitro digestion of meat and soy proteins, 2022, Food Chemistry, 383: 132465, https://doi.org/10.1016/j.foodchem.2022.132465
Richard E. Austic, 1985, Development and Adaptation of Protein Digestion, The Journal of Nutrition, 115, No 5, 686-697, https://doi.org/10.1093/jn/115.5.686
Duncan SH, Iyer A, Russell WR. Impact of protein on the composition and metabolism of the human gut microbiota and health. 2021 Proceedings of the Nutrition Society. 80, no 2 :173-185. doi:10.1017/S0029665120008022
Karr-Lilienthal, L. K., C. T. Kadzere, C. M. Grieshop, and G. C. Fahey. “Chemical and Nutritional Properties of Soybean Carbohydrates as Related to Nonruminants: A Review.” Livestock Production Science 97, no. 1 (October 1, 2005): 1–12. https://doi.org/10.1016/j.livprodsci.2005.01.015
National Animal Nutrition Program.2024. https://animalnutrition.org/feed-composition-database-signup?destination=feed-composition-database
Ibáñez, M.A., De Blas, C., Cámara, L. and Mateos, G.G., 2020. Chemical composition, protein quality and nutritive value of commercial soybean meals produced from beans from different countries: A meta-analytical study. Animal Feed Science and Technology, 267, p.114531.
Jha, R., Fouhse, J. M., Tiwari, U. P., Li, L., & Willing, B. P. (2019). Dietary fiber and intestinal health of monogastric animals. Frontiers in Veterinary Science, 6, 48.
Wang, Tong, and Lawrence A. Johnson. “Survey of Soybean Oil and Meal Qualities Produced by Different Processes.” Journal of the American Oil Chemists’ Society 78, no. 3 (March 1, 2001): 311–18. https://doi.org/10.1007/s11746-001-0262-7.
Wu, Yingzi, and Tong Wang. “Soybean Lecithin Fractionation and Functionality.” Journal of the American Oil Chemists’ Society 80, no. 4 (April 2003): 319–26. https://doi.org/10.1007/s11746-003-0697-x.
Zhu, Liuying, Wenting Li, Zeyuan Deng, Hongyan Li, and Bing Zhang. The Composition and Antioxidant Activity of Bound Phenolics in Three Legumes, and Their Metabolism and Bioaccessibility of Gastrointestinal Tract. Foods 9, no. 12 (December 2020): 1816. https://doi.org/10.3390/foods9121816.
Frontiers in Nutrition (Volume 9). Nutritional Epidemiology. Research Topics. The Role of Soy in Human Health and Disease. https://rb.gy/sw2wgs
Setchell, Kenneth DR, Nadine M. Brown, and Eva Lydeking-Olsen. "The clinical importance of the metabolite equol—a clue to the effectiveness of soy and its isoflavones." The Journal of Nutrition 132, no.12 (2002): 3577-3584.
Cassidy, A., Brown, J.E., Hawdon, A., Faughnan, M.S., King, L.J., Millward, J., Zimmer-Nechemias, L., Wolfe, B. and Setchell, K.D., 2006. Factors affecting the bioavailability of soy isoflavones in humans after ingestion of physiologically relevant levels from different soy foods. The Journal of Nutrition, 136(1), pp.45-51.
Mahfuz, Shad, Qinghui Shang, and Xiangshu Piao. Phenolic Compounds as Natural Feed Additives in Poultry and Swine Diets: A Review. Journal of Animal Science and Biotechnology 12, no. 1 (April 7, 2021): 48. https://doi.org/10.1186/s40104-021-00565-3.
Wiseman, H., J. D. O’Reilly, H. Adlercreutz, A. I. Mallet, E. A. Bowey, I. R. Rowland, and T. A. Sanders. Isoflavone Phytoestrogens Consumed in Soy Decrease F(2)-Isoprostane Concentrations and Increase Resistance of Low-Density Lipoprotein to Oxidation in Humans. The American Journal of Clinical Nutrition 72, no. 2 (August 2000): 395–400. https://doi.org/10.1093/ajcn/72.2.395.
Russell, W., and Duthie, G., Plant Secondary Metabolites and Gut Health: The Case for Phenolic Acids. Proceedings of the Nutrition Society,70(3), pp.389–396. https://doi.org/10.1017/S0029665111000152.
Ruiz-Larrea, M.B., Mohan, A.R., Paganga, G., Miller, N.J., Bolwell, G.P. and Rice-Evans, C.A., 1997. Antioxidant activity of phytoestrogenic isoflavones. Free Radical Research, 26(1), pp.63-70.
Li, Y. P., Jiang, X. R., Wei, Z. X., Cai, L., Yin, J. D., & Li, X. L. (2020). Effects of soybean isoflavones on the growth performance, intestinal morphology and antioxidative properties in pigs. Animals, 14(11), 2262-2270.
Setchell, K.D. and Clerici, C., 2010. Equol: pharmacokinetics and biological actions. The Journal of Nutrition, 140(7), pp.1363S-1368S.
Dia, V.P., Berhow, M.A. and Gonzalez De Mejia, E., 2008. Bowman−Birk inhibitor and Genistein among Soy compounds that synergistically inhibit nitric oxide and prostaglandin E2 pathways in lipopolysaccharide-induced macrophages. Journal of agricultural and food chemistry, 56(24), pp.11707-11717.
Francis, G., Z. Kerem, H.P.S. Makkar, and K. Becker. 2002. The biological action of saponins in animal systems: a review. Br. J. Nutr. 88:587–605. doi:10.1079/BJN2002725.
Kang, J.H., M.K. Sung, T. Kawada, H. Yoo, Y.K. Kim, J.S. Kim, and R. Yu. 2005. Soybean saponins suppress the release of proinflammatory mediators by LPS-stimulated peritoneal macrophages. Cancer Letters 230, no. 2 (2005): 219-227.
Zhu C, Wu Y, Jiang Z, Zheng C, Wang L, Yang X, et al. Dietary soy isoflavone attenuated growth performance and intestinal barrier functions in weaned piglets challenged with lipopolysaccharide. Int Immunopharma. 2015;28(1):288–94
Morimoto, M., Watanabe, T., Yamori, M., Takebe, M., & Wakatsuki, Y. (2009). Isoflavones regulate innate immunity and inhibit experimental colitis. Journal of Gastroenterology and Hepatology, 24(6), 1123-1129.
Andres, Aline, Sharon M. Donovan, and Mark S. Kuhlenschmidt. “Soy Isoflavones and Virus Infections.” The Journal of Nutritional Biochemistry 20, no. 8 (August 1, 2009): 563–69. https://doi.org/10.1016/j.jnutbio.2009.04.004.
Ninfali, P., Antonelli, A., Magnani, M. and Scarpa, E.S., 2020. Antiviral properties of flavonoids and delivery strategies. Nutrients, 12(9), p.2534.
Amigo, Lourdes, and Blanca Hernández-Ledesma. 2020. Current Evidence on the Bioavailability of Food Bioactive Peptides Molecules 25, no. 19: 4479. https://doi.org/10.3390/molecules25194479
Ben O. de Lumen, Lunasin: A Cancer-Preventive Soy Peptide, Nutrition Reviews, Volume 63, Issue 1, January 2005, Pages 16–21, https://doi.org/10.1111/j.1753-4887.2005.tb00106.x
Jakubczyk, Anna, Monika Karaś, Kamila Rybczyńska-Tkaczyk, Ewelina Zielińska, and Damian Zieliński. 2020. "Current Trends of Bioactive Peptides—New Sources and Therapeutic Effect" Foods 9, no. 7: 846. https://doi.org/10.3390/foods9070846
ISAPP: Gibson, G.R., Hutkins, R., Sanders, M.E., Prescott, S.L., Reimer, R.A., Salminen, S.J., Scott, K., Stanton, C., Swanson, K.S., Cani, P.D. and Verbeke, K., 2017. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology & Hepatology, 14(8), pp.491-502.
Liying, Z., Li, D., Qiao, S., Johnson, E.W., Li, B., Thacker, P.A. and Han, I.K., 2003. Effects of stachyose on performance, diarrhoea incidence and intestinal bacteria in weanling pigs. Archives of Animal Nutrition, 57(1), pp.1-10.
Nakata, T., Kyoui, D., Takahashi, H., Kimura, B. and Kuda, T., 2017. Inhibitory effects of soybean oligosaccharides and water-soluble soybean fibre on formation of putrefactive compounds from soy protein by gut microbiota. International Journal of Biological Macromolecules, 97, pp.173-180.
Zhou, X.L., Kong, X.F., Lian, G.Q., Blachier, F., Geng, M.M. and Yin, Y.L., 2014. Dietary supplementation with soybean oligosaccharides increases short-chain fatty acids but decreases protein-derived catabolites in the intestinal luminal content of weaned Huanjiang mini-piglets. Nutrition Research, 34(9), pp.780-788.
Roberfroid, M.B., 1997. Health benefits of non-digestible oligosaccharides. Dietary Fiber in Health and Disease, pp.211-219.
Liao, S.F. and Nyachoti, M., 2017. Using probiotics to improve swine gut health and nutrient utilization. Animal Nutrition, 3(4), pp.331-343.
Couto, A, T. M. Kortner, M. Penn, A. M. Bakke, Å. Krogdahl, and A. Oliva-Teles. “Effects of Dietary Phytosterols and Soy Saponins on Growth, Feed Utilization Efficiency and Intestinal Integrity of Gilthead Sea Bream (Sparus Aurata) Juveniles.” Aquaculture 432 (August 20, 2014): 295–303. https://doi.org/10.1016/j.aquaculture.2014.05.009
Castro-Gómez, P., Garcia-Serrano, A., Visioli, F. and Fontecha, J., 2015. Relevance of dietary glycerophospholipids and sphingolipids to human health. Prostaglandins, Leukotrienes and Essential Fatty Acids, 101, pp.41-51.
Treede, I., Braun, A., Sparla, R., Kuhnel, M., Giese, T., Turner, J.R., Anes, E., Kulaksiz, H., Fullekrug, J., Stremmel, W. and Griffiths, G., 2007. Anti-inflammatory effects of phosphatidylcholine. Journal of Biological Chemistry, 282(37), pp.27155-27164.
Juritsch, A.F. and Moreau, R., 2018. Role of soybean-derived bioactive compounds in inflammatory bowel disease. Nutrition Reviews, 76(8), pp.618-638.
Kurano, M., Hasegawa, K., Kunimi, M., Hara, M., Yatomi, Y., Teramoto, T. and Tsukamoto, K., 2018. Sitosterol prevents obesity-related chronic inflammation. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 1863(2), pp.191-198.