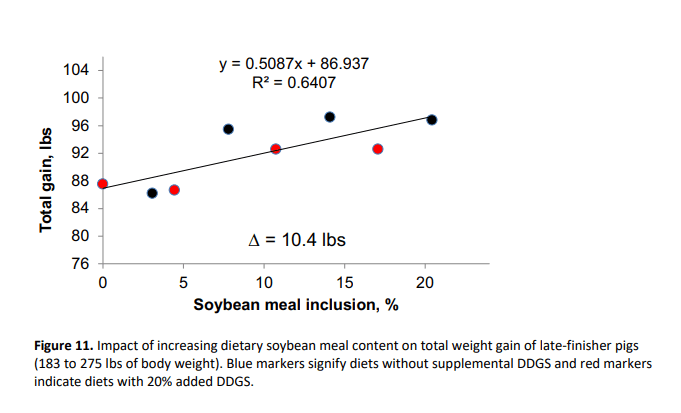Increased inclusion of soybean meal in diets improves gain and feed efficiency of healthy finishing pigs
This article is the 6th in a series highlighting the unique value of soybean meal to swine nutrition and health.
By Eric van Heugten, North Carolina State University, Raleigh, NC
Introduction
Soybean meal (SBM) is an excellent protein source for pigs because it has a well-balanced amino acid profile, providing high concentrations of both essential and nonessential amino acids. This makes SBM very cost-competitive compared to other protein sources (Ibáñez et al., 2020; Pope et al., 2023). The energy concentration in SBM positively affects its utility in practical diets for swine, with recent studies indicating that the net energy level in SBM is greater than previously reported, further increasing the value proposition of SBM (Cemin et al., 2020; Lee et al., 2022; Pope et al., 2023; Boyd and Gaines, 2023). However, inclusion levels of SBM in swine diets have decreased over time, mainly because of the increased availability of crystalline amino acids, the use of distillers’ dried grains with solubles (DDGS), and the price reduction of tryptophan; the latter causing dietary SBM content to dramatically decrease. For example, the average diets in the year 2000 did not contain DDGS and included approximately 3 lbs/ton of L-lysine·HCl. In 2008, the inclusion of up to 12 lbs/ton of lysine became possible because tryptophan became affordable. In 2010, DDGS started to be included at approximately 20% with the addition of crystalline amino acids. In 2017, DDGS were included at levels well over 20% with high levels of crystalline amino acids. During this timeframe (2000, 2008, 2010, 2017), diets contained approximately 464, 346, 172, and 52 lbs per ton of SBM, respectively. With crystalline tryptophan being reliably cost-effective, and now valine and isoleucine also available at competitive costs, there is greater potential to drive dietary SBM content even lower, possibly down to zero in pigs weighing about 160 lbs or more.
Thus, the inclusion of SBM in pig diets has become largely dependent on pricing, which does not account for the potential value of SBM beyond its contribution as a source of amino acids. In addition to its value as a protein and energy source, SBM contains significant amounts of health-promoting compounds, including isoflavones, saponins, and phenolic antioxidants (Petry et al., 2024; Sharma et al., 2019; Smith and Dilger, 2018), as well as bioactive peptides generated from the digestion of soy proteins (Chatterjee et al., 2018). Several studies have shown benefits of increased SBM inclusion in diets of pigs exposed to respiratory diseases, most notably porcine reproductive and respiratory syndrome (Boyd et al., 2010; Zier-Rush et al., 2015; Rochel et al., 2015). Thus, SBM can be strategically used at higher levels to mitigate the negative impacts of respiratory disease on growth performance, feed efficiency, medication costs, and full value pigs (Boyd et al., 2023). Functional components in SBM may also provide benefits in healthy pigs as suggested by improved daily gain and feed efficiency, as well as reduced medical treatments and culling with increasing dietary levels of SBM for late nursery pigs (Moran et al., 2017; Cemin et al., 2019, 2020). Price competitiveness of DDGS and crystalline amino acids have forced dietary SBM inclusion to very low levels, which may reduce possible benefits from functional components in SBM that take it beyond serving as a well-balanced source of amino acids and energy.
Purpose of this study
To evaluate the potential benefits of SBM in healthy pigs, two studies were conducted using early- and late-finishing pigs to determine the impact of SBM displacement on the growth performance of pigs. We hypothesized that extensive replacement of SBM from the diet would impair growth performance, even in healthy pigs. The objectives were to determine the impact of SBM displacement with DDGS and crystalline amino acids on growth performance and carcass quality of healthy early- and late-finishing pigs in two separate studies conducted sequentially (adapted from Anderson, 2021). We further aimed to construct response curves to dietary SBM inclusion based on growth phase to account for potential difference in response due to pig body weight.
Experimental design of studies
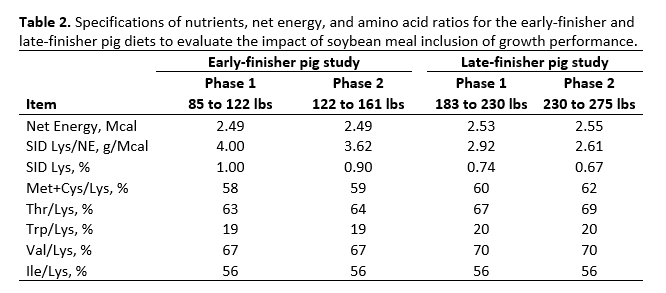
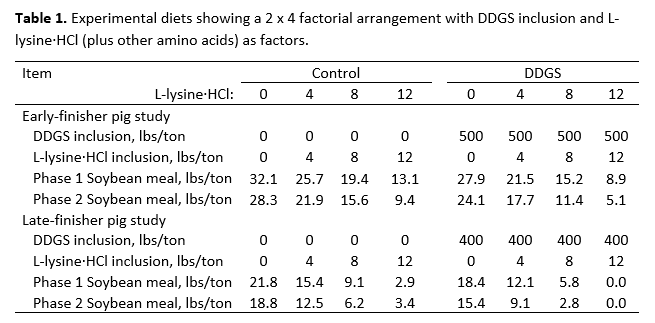
Pigs were assigned to one of 8 dietary treatments arranged in a 2 x 4 factorial (Table 1) with 2 levels of DDGS (0 or 25% for the early-finisher pig study and 0 or 20% for the late-finisher pig study) and 4 levels of crystalline amino acid supplementation. In the early-finisher pig study, a total of 512 pigs with an average starting body weight of 84.9 ± 0.3 lbs were grouped by body weight and sex and assigned to 64 pens with four gilts and four barrows per pen, resulting in eight replicates and 64 pigs per treatment. In the late-finisher pig study, a total of 480 pigs with an average starting body weight of 183.2 ± 0.8 lbs were grouped by body weight and sex and assigned to 80 pens with three gilts and three barrows per pen, resulting in 10 replicates and 60 pigs per treatment. In both studies, amino acid supplementation consisted of L-lysine HCl added at 0, 4, 8, or 12 lbs per ton, with DL-methionine, L-threonine, L-tryptophan, L-valine, and L-isoleucine supplemented to maintain appropriate standardized ileal digestible (SID) amino acid ratios (Table 2). The ratios used were obtained from van der Peet-Schwering and Bikker (2018) and exceeded NRC (2012) recommendations.
Diet formulation
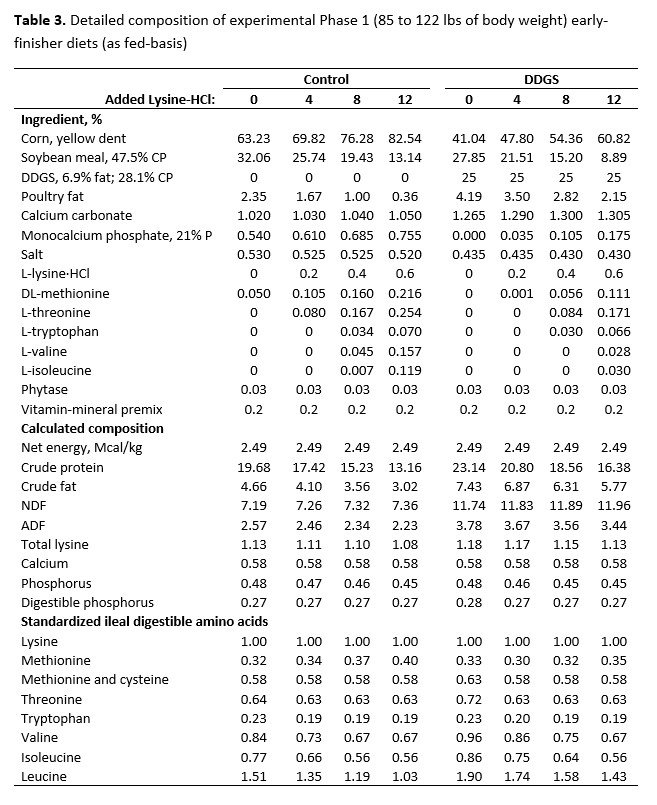
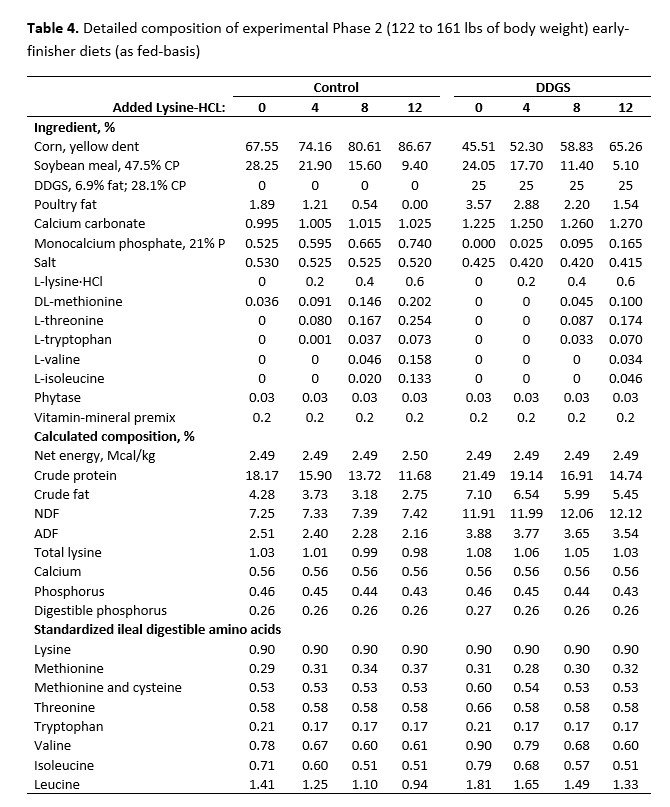
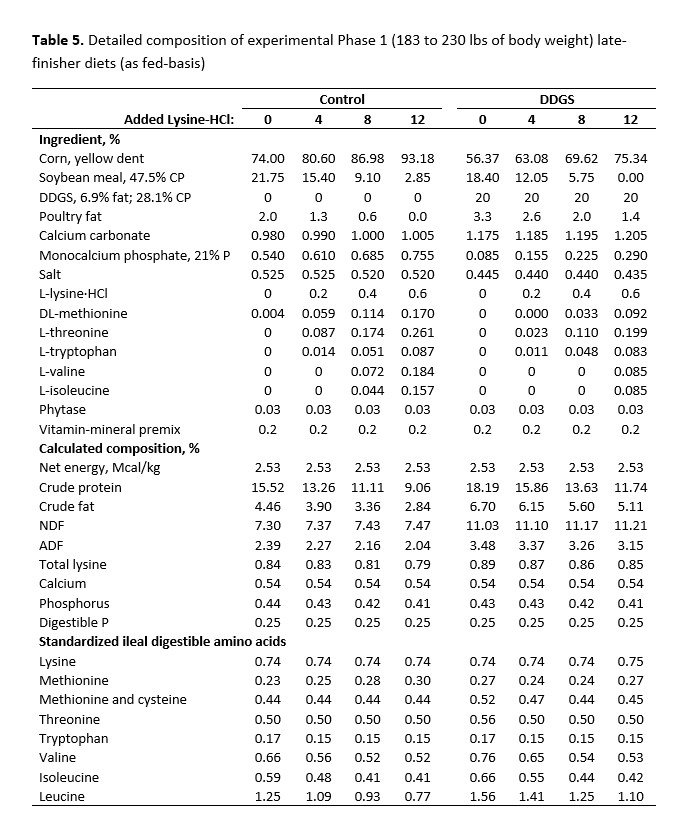
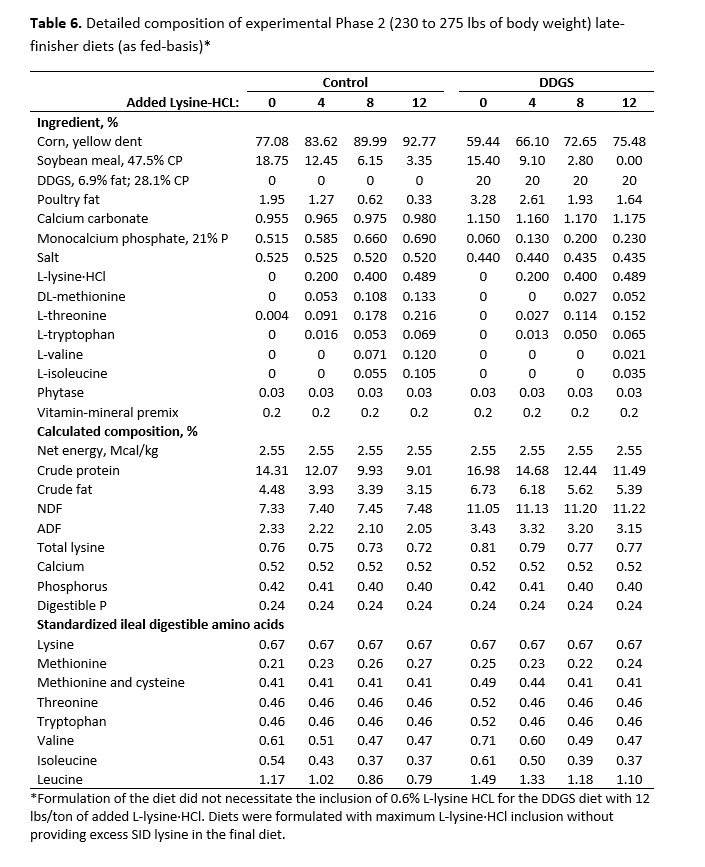
For both studies, dietary treatments were formulated by a commercial feed company to take advantage of their nutrient matrix values for the ingredients used and to ensure these values were as current and accurate as possible. The net energy value used for SBM in the diet formulation was more conservative than the more recent net energy value suggested by Boyd and Gaines (2023). For the early-finisher study, diets were fed in two phases and were formulated to contain 1.00% (Phase 1, 21 days) and 0.90% (Phase 2, 18 days) standardized ileal digestible (SID) lysine, and were carefully balanced for ideal protein and net energy. Dietary SBM inclusion declined with the inclusion of crystalline amino acids from 32.06% to 13.14% (Phase 1) and 28.25% to 9.40% (Phase 2) for control diets without DDGS (Table 1). Inclusion of SBM declined from 27.85% to 8.89% (Phase 1) and 24.05% to 5.10% (Phase 2) with increasing crystalline amino acid inclusion for diets with DDGS.
For the late-finisher study, diets were formulated to contain 0.74% and 0.67% for Phase 1 (21 days) and Phase 2 (13 days for the heaviest five replicates and 20 days for the lightest five replicates) diets, respectively. Diets were balanced for SID lysine, methionine and cysteine, threonine, tryptophan, valine, and isoleucine and were equal in net energy. Inclusion of SBM decreased with increasing levels of crystalline amino acids from 21.75% to 2.85% for Phase 1 and from 18.75% to 3.35% for Phase 2 for the control diets. It decreased from 18.40% to 0% for Phase 1 and 15.40 to 0% for diets with DDGS. For Phase 2 diets with DDGS, only 0.427% added L-lysine HCl was required to meet SID lysine requirements for the highest crystalline amino acids supplemented diet. Thus, crystalline L-lysine·HCl supplementation was limited to a maximum of 8.54 lbs/ton for Phase 2 diets. Diet composition for Phase 1 and Phase 2 diets for both early- and late-finisher studies are shown in Tables 3 to 6. All diets were chemically verified and were consistent with the targeted formulated values.
Benefits of SBM in early-finishing pigs
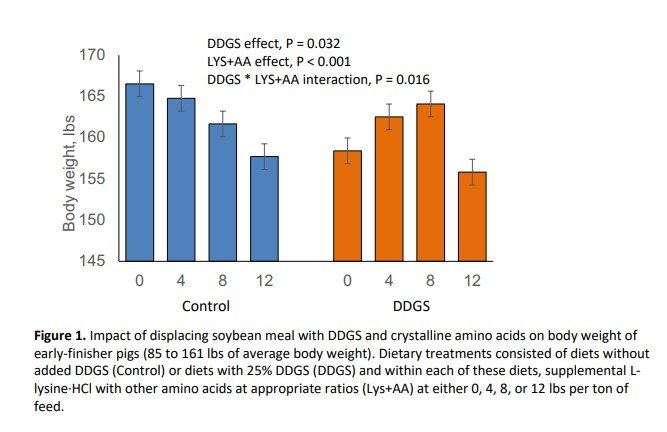
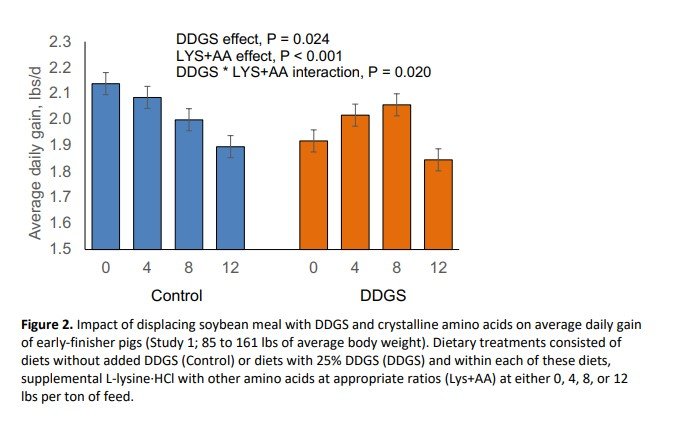
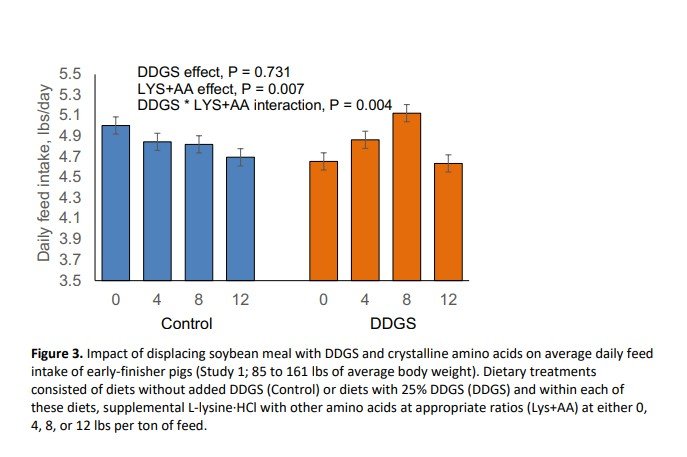
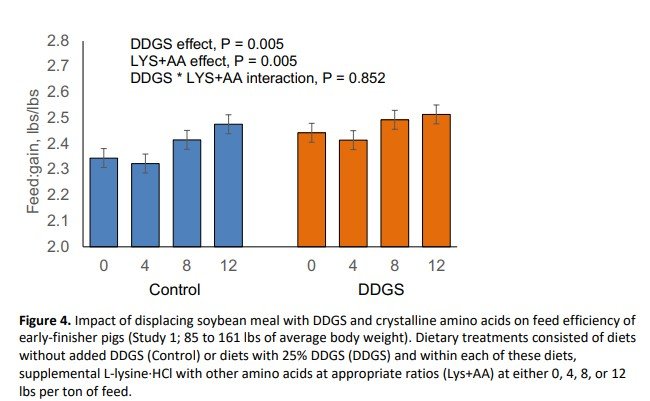
Inclusion of DDGS at 25% in partial replacement of SBM reduced final body weight (P = 0.032; Figure 1) and average daily gain (P = 0.024; Figure 2) during the 39-day experimental period. Supplementation with crystalline amino acids reduced (P < 0.001) final body weight and average daily gain (Figures 1 and 2). Pigs fed the DDGS diet had a 2.47 lbs lower body weight at the end of the study, whereas replacing SBM with 12 lbs/ton of L-lysine·HCl (plus other crystalline amino acids) reduced final body weight by 5.71 lbs. The response to supplemental crystalline amino acids was dependent on whether DDGS were included or not. Supplementation of crystalline amino acids decreased growth rate and final body weight linearly when DDGS were not included in the diet, whereas the decrease was quadratic when DDGS were included. Feed intake was not impacted (P = 0.731) by DDGS supplementation (Figure 3). However, feed intake linearly decreased with crystalline amino acid supplementation within control diets, but in a quadratic manner when supplemented to DDGS-containing diets (interaction, P = 0.004; Figure 3). Feed efficiency expressed as feed:gain was 3.2% higher (P = 0.005) for pigs fed diets with DDGS compared to control-fed pigs, consuming 0.08 lbs more feed per lb of gain (Figure 4). Feeding diets with supplemental crystalline amino acids (12 lbs/ton of L-lysine·HCl) resulted in a 4.2% higher feed:gain (P = 0.005) or 0.10 lbs more feed per lb of gain.
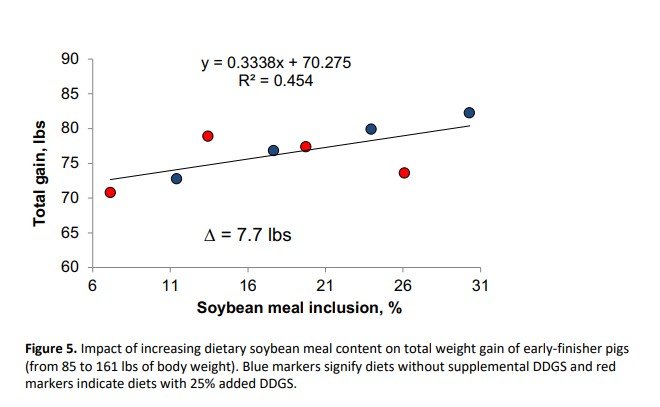
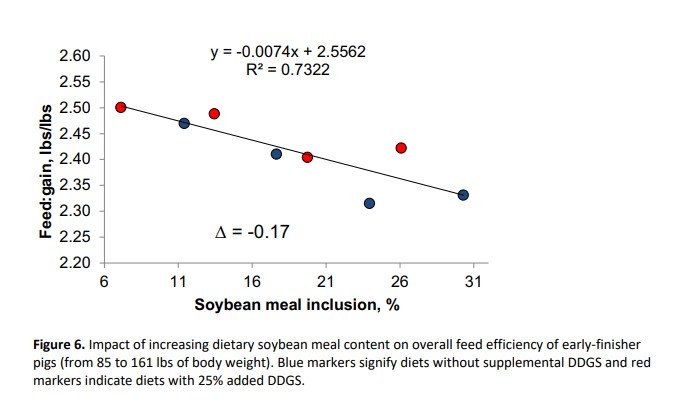
This study showed that displacement of SBM with DDGS reduced growth rate and increased feed:gain, whereas supplementation with crystalline amino acids negatively affected growth and feed efficiency for SBM control diets, but did not cause further reductions in DDGS-containing diets. Diets in this study were carefully formulated to meet requirements for standardized ileal digestible lysine, methionine plus cysteine, threonine, tryptophan, valine and isoleucine, yet growth performance was compromised with high inclusion of crystalline amino acids. Thus, increasing the amount of SBM in the diet was positively correlated with improved total weight gain (R2 = 0.454) and feed efficiency (R2 = 0.732) as can be seen in Figures 5 and 6, respectively. The response of total weight gain to SBM inclusion was more evident within the control diets (blue circles) than the DDGS-containing diets (red circles). On the other hand, feed efficiency linearly improved with increasing inclusion of SBM regardless of DDGS inclusion.
The regression analysis indicated that for every 1% increase in SBM inclusion, there was a 0.33 lbs increase in total gain and an improvement in feed efficiency of 0.007. This study was conducted during the winter with high health status pigs, and other work (Boyd et al., 2010; Zier-Rush et al., 2015; Rochel et al., 2015) suggests that the impact of dietary SBM would be even greater in pigs exposed to heat stress or respiratory health challenges.
Benefits of SBM in late-finishing pigs
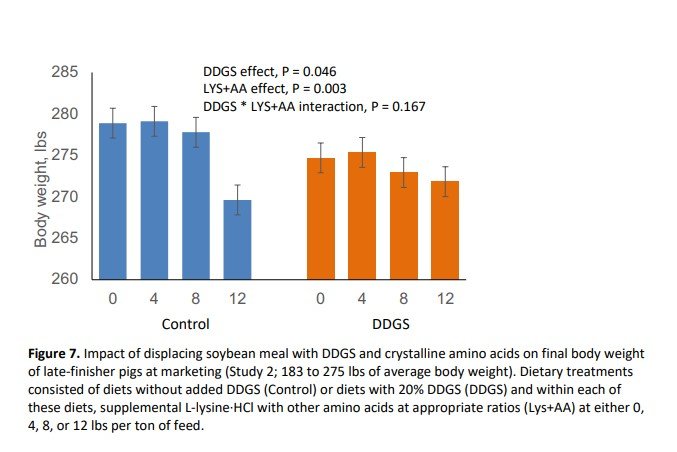
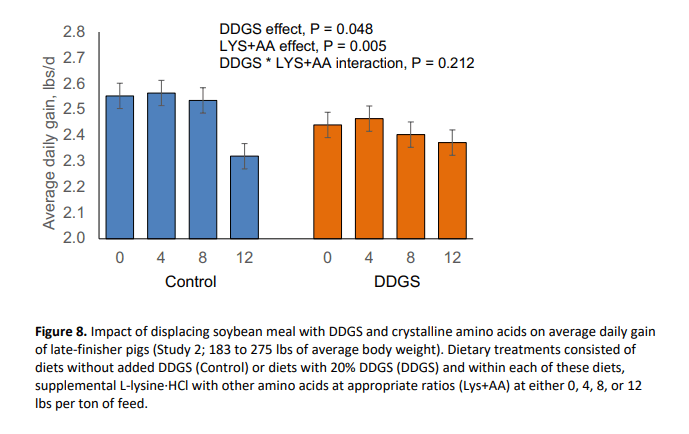
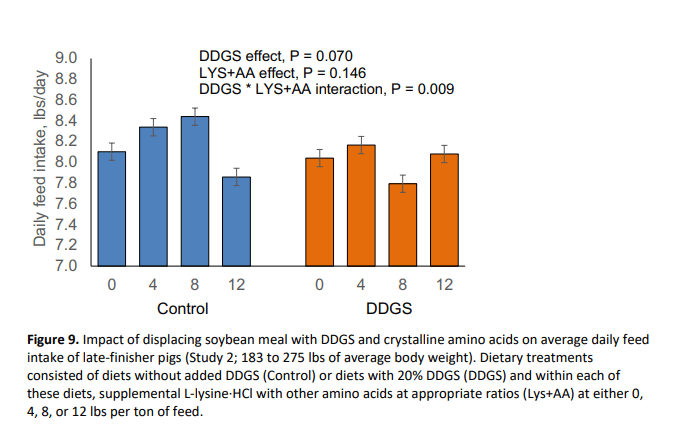
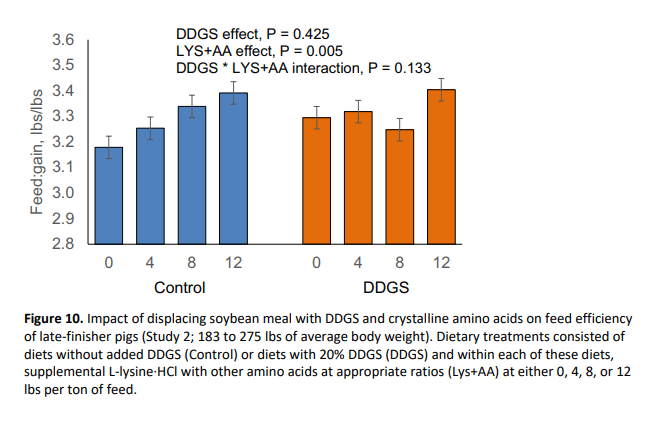
In this study, pig body weight at marketing was reduced by 2.69 lbs (P = 0.046) with the inclusion of DDGS (Figure 7). Further, DDGS inclusion reduced (P = 0.048) average daily gain (Figure 8) and tended to decrease daily feed intake (P = 0.070; Figure 9) without affecting feed efficiency (Figure 10). Supplemental crystalline amino acids linearly decreased (P < 0.005) body weight and average daily gain, without impacting feed intake, and worsened feed efficiency (P = 0.005). Inclusion of 12 lbs/ton of L-lysine·HCl (8.54 lbs/ton for Phase 2) while balancing other essential amino acids decreased final market weight by 6.06 lbs and increased feed:gain by 5.0% which is equivalent to 0.162 lbs of extra feed per lb of gain.
Ultrasound back fat thickness tended to be reduced (0.60 vs. 0.62 in.) when DDGS were included (P = 0.071), and crystalline amino acids quadratically influenced loin eye area (7.51, 7.58, 7.59, and 7.33 in2; P = 0.011). These apparent differences in ultrasound carcass measurements were directly related to the decreased final body weight in pigs fed DDGS and crystalline amino acids. Carcass yield was reduced by an absolute 0.42% (72.62 vs. 73.04) with DDGS supplementation (P = 0.034). Therefore, marginal carcass weight was 3.08 lbs lower with DDGS inclusion (data not shown).
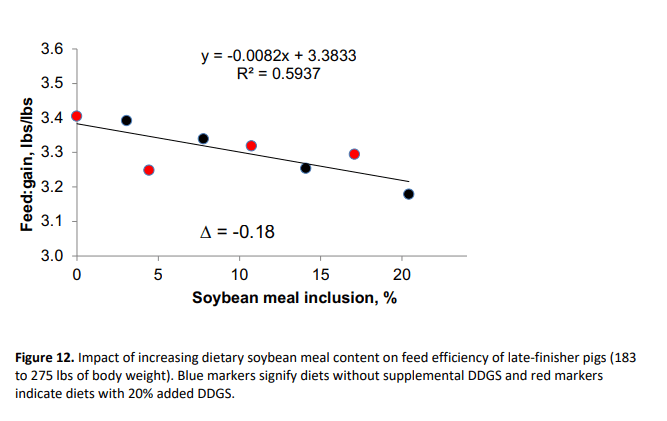
Collectively, these data indicate that displacement of SBM with DDGS and crystalline amino acids, especially at the highest level of crystalline amino acid supplementation, negatively affected growth of late-finishing pigs. In other words, increasing the amount of dietary SBM improved total weight gain (R2 of 0.64) and feed efficiency (R2 of 0.59) as shown in Figures 11 and 12, respectively. This response suggests that for every 1% increase in SBM inclusion, total gain was improved by 0.51 lbs and feed:gain was reduced by 0.008. It should be noted that pigs in the present study were extremely healthy and that the effects of dietary SBM are expected to be more pronounced under respiratory health challenges (Boyd et al., 2010; Zier-Rush et al., 2015; Rochel et al., 2015). The impacts observed here would likely be of greater importance especially in commercial production during the summer when average daily gain is at a premium to achieve target market weights.
Practical application of SBM in diets for finishing pigs
In both studies described here, the displacement of SBM by DDGS and crystalline amino acids negatively impacted both body weight gain and feed efficiency. Increasing the level of dietary SBM had a remarkable impact on maximizing gain. Comparing the highest to the lowest inclusion level of SBM on live weight gain yielded approximately 18 lbs of extra live body weight for both the early-finisher and the late-finisher study combined. Marginal feed efficiency, from the lowest to the highest SBM inclusion level improved by 0.17 and 0.23, for the early-finisher and late-finisher, respectively. In a fixed-time scenario and especially when pigs are profitable, maximum gain is valuable, whereas strictly focusing on feed cost will compromise return over investment.
Part of the observed response was due to the negative impact of DDGS on growth performance in both early- and late-finishing pigs. Shurson (2019) summarized results of DDGS inclusion in diets for finishing pigs for studies conducted in 2010 or later. The calculated reduction in average daily gain in that review was moderate and averaged 0.91%, 3.20%, and 2.44% with the inclusion of DDGS at 15% to 20%, 25% to 30%, and more than 30%, respectively. A reduction in carcass yield is relatively common when feeding DDGS because of its high fiber content, which reduces carcass gain and also reduces carcass gain relative to feed intake, which is consistent with the data presented herein.
It should be noted that these previously published studies did not use the extraordinary levels of amino acids that we use today, and many of the early studies with DDGS were conducted when tryptophan was expensive. Without the price competitive availability of tryptophan, the inclusion level of SBM would have been reduced to a much lesser extent with the inclusion of DDGS in these earlier studies. In today’s market, with the availability of tryptophan, valine and even isoleucine, there is much more pressure for exact application of proper amino acid ratios in diet formulation than in the past. In addition, with the availability of tryptophan and other amino acids, the inclusion of SBM drops substantially when DDGS are included. If SBM has growth-stimulating properties (Petry et al., 2024), or if amino acids beyond tryptophan (and valine and isoleucine) become limiting due to the reduction in SBM inclusion, then compromised performance of pigs should be expected when dietary SBM is reduced.
In the studies described here, replacement of SBM was carefully balanced through formulation of diets that met or exceeded requirements for SID lysine, methionine plus cysteine, threonine, tryptophan, valine and isoleucine and were equal in net energy content. When formulating diets with high levels of crystalline amino acids, total crude protein in the diet decreased. In our studies, this reduction in crude protein was rather drastic. In general terms, research suggests that protein can be reduced by 2.5% to 3%, but beyond that point, growth and feed efficiency is generally impaired (Boyd et al., 2024). Johnson, Usry, and Boyd reported a study in 1999 that showed that as SBM content decreased from 375 lbs/ton to near zero (in 110 lb pigs), pigs became fatter and less efficient despite having restored the original dietary balance of all indispensable and dispensable amino acids (Boyd et al., 2024). Potassium level and dietary electrolyte balance was also maintained equal to or above a safe minimum. They proposed that the observed responses were not due to a crude protein limitation, but rather were related to growth and health-promoting components in SBM that is not common to alternative protein sources. It is interesting to note that practical, applied studies evaluating the minimum protein requirements of pigs often reduce dietary SBM while supplementing with high levels of crystalline amino acids to incrementally reduce dietary protein. In this case, effects of a deficiency in non-essential amino acids with reduced crude protein diets cannot be clearly distinguished from potential effects of health-promoting molecules from SBM (Soto et al., 2019).
Does dietary protein minimum make sense?
Suggesting a minimum requirement of crude protein does not provide much accuracy, considering that in diets with relatively poor protein sources crude protein will be unnecessarily high and should be able to be reduced to a greater extent than when high-quality protein sources are used. The premise that non-essential amino acids may be limiting growth in pigs fed diets that are low in crude protein, but meet the requirements for all essential amino acids is valid, but perhaps needs updating. Recent studies suggest that under certain circumstances pigs may be incapable of synthesizing adequate quantities of certain non-essential amino acids to fully express their genetic potential (Wu et al., 2014; Hou et al., 2015; Wu and Li, 2022). Under these conditions, growth or other physiological processes may be compromised unless these conditionally essential amino acids are provided at sufficient quantities in the diet. The possibility of synthesizable amino acids (either conditionally essential or non-essential amino acids) playing a role in the observed responses in these studies cannot be ruled out. If this is the case, increased levels of SBM were able to correct this deficiency.
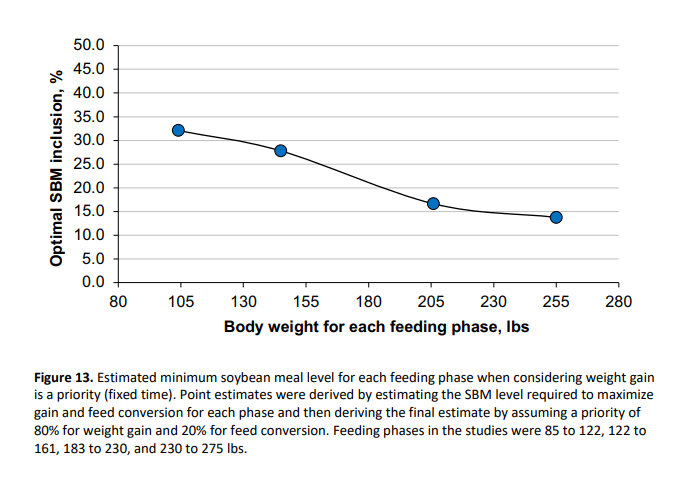
To enhance translation of these research findings into practical application, we estimated the approximate minimum SBM inclusion level for each feeding phase that would maximize gain. This serves as a starting point for recommendations for minimum SBM inclusion at various body weights of finishing pigs (Figure 13). Feed conversion was also improved in our studies, but the level of SBM required to optimize it was slightly different. The minimum SBM levels shown in Figure 13 were derived mainly for maximum growth, but also account partly for improved feed efficiency, and they assume fixed time conditions (e.g., summer). Fastest growth tends to produce the highest percentage of full-value price market pigs and the least cull-value market pigs in the population. In practice, this proposed curve should be validated and refined to derive the optimum population profit outcome. Financial optimization integrates feed input cost against value created; however, when pigs are profitable, and time is fixed, pounds of full-value pigs produced overwhelmingly drive the SBM specification. Further calibration of SBM inclusion must be based on population outcomes, and must make the specifications applicable to the specific production system and time of year. Answers are expected to be different for Summer versus non-summer, which needs to be validated. Addition of a normal summer diet as a control when developing a summer SBM optimum curve will be crucial in order to verify whether maximizing growth by preventing SBM replacement from being too extreme is beneficial under those conditions.
Key Conclusions
In early-finishing pigs, displacement of SBM from 31% to 6% in growing pig diets compromised weight gain by over 7 lbs and feed:gain ratio by 0.17 units.
In late-finishing pigs, displacement of SBM from 21% to 0% compromised weight gain by 10 lbs and feed:gain ratio by 0.18 units.
Minimum SBM specifications throughout the growth cycle of finishing pigs can maximize profitability, especially in a fixed-time scenario.
Biologically functional components other than amino acids present in SBM play a role in the improvements in growth performance, even in healthy pig populations.
References
Ibáñez, M.A., C. de Blas, L. Cámara, and G.G. Mateos. 2020. Chemical composition, protein quality and nutritive value of commercial soybean meals produced from beans from different countries: A meta-analytical study. Anim. Feed Sci. Techn. 267:114531.
Pope, M., B. Borg, R.D. Boyd, D. Holzgraefe, C. Rush, and M. Sifri. 2023. Quantifying the value of soybean meal in poultry and swine diets. J. Appl. Poultry Res. 32:100337.
Boyd, R.D. and A.M. Gaines. 2023. Soybean meal NE value for growing pigs is greater in commercial environments. Feedstuffs, August digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-august-2023/page/2/1.
Cemin, H. S., H. E. Williams, M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. Derouchey, K. F. Coble, B. A. Carrender, and M. J. Gerhart. 2020. Estimate of the energy value of soybean meal relative to corn based on growth performance of nursery pigs. J. Anim. Sci. Biotech 11:70. https://doi.org/10.1186/s40104-020-00474-x.
Lee, S.A., D.A. Rodriguez and H.H. Stein. 2022. Net energy in U.S. soybean meal fed to group-housed growing pigs is greater than calculated book values. 15th Int. Symp. Diges. Physio. Pigs. Animal, Sci. proc. 13 (Issue 2):178.
Petry, A. L., B. Bowen, L. Weaver, and R. D. Boyd. 2024. Functional compounds in soybean meal: implications for pig health and physiology. Feedstuffs, February digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-february-2024/page/5/1
Sharma, R., S. Kumar, V. Kumar, and A. Thakur. 2019. Comprehensive review on nutraceutical significance of phytochemicals as functional food ingredients for human health management. J. Pharma. Phytochem. 8 (5):385-395. http://doi.org/10.22271/phyto.2019.v8.i5h.9589.
Smith, B. S., and R. N. Dilger. 2018. Immunomodulatory potential of dietary soybean-derived isoflavones and saponins in pigs. J. Anim. Sci. 96:1288–1304. https://doi.org/10.1093/jas/sky036.
Chatterjee, C., S. Gleddie and C-W Xiao. 2018. Soybean bioactive peptides and their functional properties: Review. Nutrients 10 (9):1211-1227. http://dx.doi.org/1.3390/nu10091211.
Boyd R. D., M.E. Johnston, and C.E. Zier-Rush. 2010. Soybean meal level modulates the adverse effect of high immune stress on growth and feed efficiency in growing pigs. In: Proc 71st Minnesota Nutrition Conf, Owatonna, MN: University of Minnesota, St. Paul, MN.
Zier-Rush, C., S. Smith, J. Picou, E. van Heugten and R.D. Boyd. 2015. High SBM level reduces medical treatment of PRRS affected weaned pigs fed early nursery diets. Hanor Tech. Memo H-2015-07. https://doi.org/10.13140/RG.2.2.35498.39366.
Rochell S.J., L.S. Alexander, G.C. Rocha, W.G. Van Alstine, R.D. Boyd, J.E Pettigrew, and R.N. Dilger. 2015. Effects of dietary soybean meal concentration on growth and immune response of pigs infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. 93:2987–2997. doi:10.2527/jas.2014-8462.
Boyd, R.D., M. Johnston, J. Usry, P. Yeske, and A. Gaines. 2023. Soybean meal mitigates respiratory disease-impaired growth in pigs. Feedstuffs, October digital edition, page 1. https://informamarkets.turtl.co/story/feedstuffs-october-2023/page/2/1.
Moran K, R.D. Boyd, C. Zier-Rush, P. Wilcock, N. Bajjalieh, and E. van Heugten. 2017. Effects of high inclusion of soybean meal and a phytase superdose on growth performance of weaned pigs housed under the rigors of commercial conditions. J. Anim. Sci. 95:5455-65.
Cemin H.S., M.D. Tokach, A.M. Gaines, B.W. Ratliff, S.S. Dritz, J.C. Woodworth, and R. Goodband. 2019. Effects of soybean meal level and distillers dried grains inclusion on growth performance of late nursery pigs. J. Anim. Sci. 97(E-Suppl. 2):88-89(Abstr.).
Anderson, B.E. 2021. Impact on performance and carcass characteristics when replacing soybean meal with distillers dried grains with solubles and crystalline amino acids in diets of growing and finishing pigs. North Carolina State University Thesis. https://repository.lib.ncsu.edu/server/api/core/bitstreams/d5fc903e-e57b-4e49-979b-cf584b8b1d55/content
van der Peet-Schwering, C.M.C., and P. Bikker, 2018. Amino acid requirement of growing and finishing pigs. Wageningen Livestock Research, Report 1101.
National Research Council (NRC). 2012. Nutrient requirements of swine 11th revised edition. National Academy Press, Washington, DC, USA.
Shurson, G.C. 2019. U.S. Grains Council DDGS User’s Handbook. U.S. Grains Council, Washington, DC.
Boyd, R.D., M. Johnston, J. Usry, and R.E. Austic. 2024. Foundation Study for Low Protein Diet Research: SBM depletion impaired growth in pigs despite restoration of all essential amino acids and non-essential amino acid nitrogen. Feedstuffs, April digital edition. https://informamarkets.turtl.co/story/feedstuffs-april-2024/page/1.
Soto, J.A., M.D. Tokach, S.S. Dritz, J.C. Woodworth, J.M. DeRouchey, R.D. Goodband, and F. Wu. 2019. Optimal dietary standardized ileal digestible lysine and crude protein concentration for growth and carcass performance in finishing pigs weighing greater than 100 kg. J. Anim. Sci. 97(4): 1701-1711. https://doi.org/10.1093/jas/skz052.
Wu, G. 2014. Dietary requirements of synthesizable amino acids by animals: a paradigm shift in protein nutrition. J. Anim. Sci. Biotechn. 5, Article number: 34.
Hou, Y., Y. Yin, and G. Wu. 2015. Dietary essentiality of ‘‘nutritionally non-essential amino acids’’ for animals and humans. Exp. Biol. Med. 240: 997-1007. DOI: 10.1177/1535370215587913.
Wu, G. and P. Li. 2022. The “ideal protein” concept is not ideal in animal nutrition. Exp. Biol. Med. Vol. 247:1191-1201. https://doi.org/10.1177/15353702221082658.
Eric van Heugten, PhD is Professor of Nutrition and Swine Extension Specialist in the Department of Animal Science at North Carolina State University, Raleigh, NC.
Acknowledgments:
This research was funded in part by the United Soybean Board, St. Louis, MO. Data presented were adapted from the MS thesis of Brooke Anderson (North Carolina State University). Results were also presented in part at the John F. Patience International symposium on swine nutrition, August 2021.