Soybean meal NE value for growing pigs is greater in commercial environments
This is Part 2 of a 2 Part series defining the amino acid and energy specifications of soybean meal for growing pigs and poultry.
By R. Dean Boyd and Aaron M. Gaines Introduction
It is proving difficult to establish reliable net energy (NE) values for soybean meal (SBM). In a previous article, we presented NE estimates that we consider to be the minimum for growing pigs (Boyd et al., 2023). They are based on recent animal growth and calorimetry assays and exceed values provided by 4 international ingredient composition tables by 8-15%. There also appears to be a difference in SBM NE values determined in academic facilities with strict environmental control and those derived in commercial settings. Academic facilities lack pathogen (type, density) and (or) environmental stressors that are encountered in commercial barns housing 1000 or more pigs per room. We believe that this apparent conflict in NE estimates between environments is explainable by the health-promoting molecules of SBM.
Feed composition tables rely on equations that predict SBM NE from nutritional composition, but underestimate SBM NE for growing pigs when compared to recent empirical results (Lee et al., 2022). However, if SBM NE is predicted from the starting point of digestible energy (DE), it aligns better with results from calorimetry and growth assays. DE is cumbersome but this step addresses the question of energy substrate digestibility and accounts for energy components that are not included in some prediction equations. The unexpectedly high estimates for SBM NE that were obtained for the commercial setting adds to the conundrum of SBM NE value (Boyd and Rush, 2019).
In order to determine whether SBM NE estimates differ when determined in an academic setting as compared to a commercial environment, we established a baseline to represent the former using publications where DE was the starting point and one with indirect calorimetry (pens of pigs fed ad libitum). Probing for a difference between environments required reliable empirical estimates of SBM NE from the academic sector. Upon concluding that there is a difference, we propose that nutritionists use higher SBM NE values that better reflect commercial conditions. Our conceptual basis for this difference is based on the level of immune stress imposed by the environment. In this framework, pathogens divert absorbed energy to survival functions, but health-promoting molecules may prevent total diet NE diversion by intervening to minimize pathogen triggering of the immune system. This hypothesis had its origin with the study by Boyd and co-workers (2010).
Reliable SBM NE values from academic environments
The question of whether SBM NE estimates differ between a highly controlled academic (or equivalent) setting and a commercial environment requires reliable NE estimates for both. It is clear that SBM NE estimates from major international references, including the NRC (2012), are not reliable. A number of publications have appeared since the NRC was published where SBM NE was estimated from empirical studies in the University setting. In the commercial sector, the growth assay method is used to estimate SBM NE. We expect the growth assay estimate for SBM NE to be in agreement with reliable academic estimates unless immune or other stressors in the commercial environment are a significant factor. The basis for this is discussed later.
In order to make a comparison of SBM NE estimates between the two environments, we established a more reliable SBM NE estimate from the academic setting using recent publications from the Hans Stein lab (University of Illinois). We selected publications that used growing pigs (>50 lbs) and met two criteria:
SBM DE was determined as the starting point for computing NE,
Corn DE was determined as an NE reference.
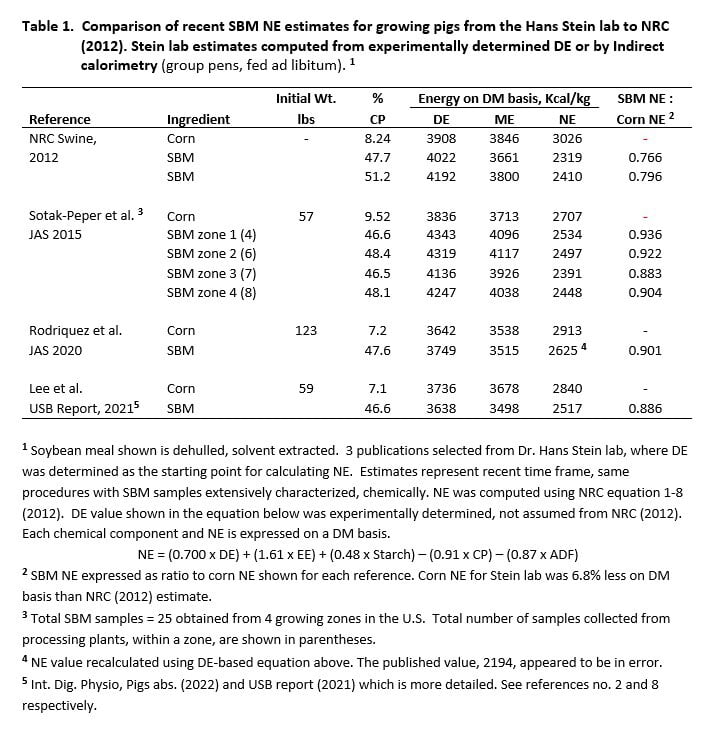
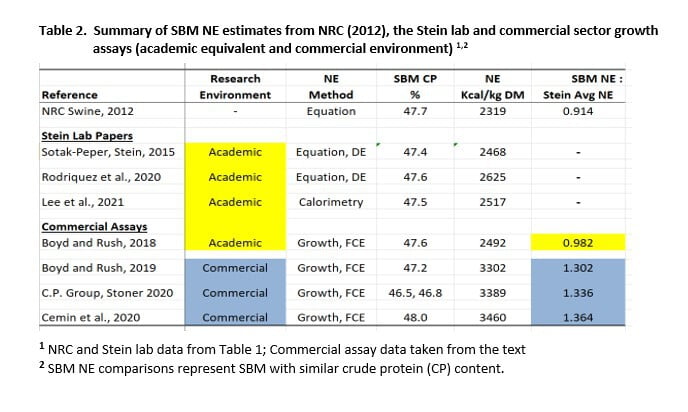
Three of 11 papers published from 2009 to 2021 met these criteria. One of the Stein lab studies involved indirect calorimetry (group pens, fed ad libitum), which is the gold standard for NE determination. A comparison of SBM NE estimates from the Stein lab and the NRC (2012) is shown in Table 1. The SBM NE to corn NE ratio was also computed for additional reference when comparing SBM NE values for academic and commercial study environments.
SBM NE values from the Stein lab exceeded the NRC (2012) estimate for crude protein (CP) equivalent SBM (47.7%) by 183 kcal/kg DM and exceeded Brazil feed composition estimates by more than 200 kcal (Rostagno et al., 2017). Comparisons in Table 1 involved a narrow range in SBM CP (46.5-48.4%) which is important since NE increases as SBM CP increases (Boyd et al., 2023). The disparity between the Stein lab SBM NE and NRC (2012) is further illustrated by SBM NE ranging from 88.3 to 93.6% of assay corn NE, while NRC estimates ranged from 76.6 to 79.6% of the respective corn NE standard.
We conclude that SBM NE is approximately 2502 kcal/kg DM for the academic environment, based on empirical estimates from the Stein lab (Table 1).
Snyder growth assay method of ingredient NE verification
The growth assay is based on relating the energetic value of a test ingredient (unknown) to that of the corn (known) it replaces. It is based in the FCE response (feed conversion efficiency) to incremental changes in the amount of a test ingredient. FCE is a discriminating measure of how diet NE is affected by substituting the test ingredient for a carefully defined corn standard. We described this procedure as the Snyder growth assay (Boyd et al., 2010). A detailed procedural document is publicly available (Boyd and Rush, 2017). Indispensable elements of the Snyder growth assay method are provided in a Backgrounder within this document. This method of validating ingredient NE values has been used by at least 2 nutrition companies for 35 years, but was kept confidential.
The growth assay has proven to be reliable for validating ingredient NE values, but SBM may present a special case since it is an abundant source of health-promoting molecules. It is conceivable that these non-nutritive components might influence how efficiently total diet NE is used for growth. The assay is structured to reflect classic ingredient NE, but the estimate would be greater if a non-nutritive fraction caused total diet NE to be used more efficiently. This nuance has not been observed with ingredients, but SBM may be the exception; an effect that would not be expressed except in the commercial environment.
Hanor studies on SBM NE have confirmed the academic estimate while also observing a higher NE value in the commercial environment; the outcome being dependent on the testing environment. These studies are described below.
First observation that SBM NE is greater in commercial environment
The first indication that SBM NE is greater for pigs in the commercial environment than the academic setting arose while conducting SBM NE calibration studies at the Hanor company. We use the growth assay in commercial barns (retrofitted for research) to confirm NE values for major ingredients.
Our first SBM NE assay was conducted in a small research facility that Hanor used to test concepts before moving to the scale and complexity of a commercial facility. The experimental setting involved small group size (24 pens, 10 mixed sex pigs/pen) and good environmental conditions (temperature, population density), similar to an academic setting (Boyd and Rush, 2018). Pigs were sourced from a sow farm that was positive for PRRSv, but stable. They appeared healthy during the test with no clinical signs of disease. The growth assay was conducted for 28 d with pigs over the 101 to 165 lbs phase of growth. This research was prompted by the wide range of published estimates of SBM NE.
A second SBM growth assay was conducted at the Hanor commercial research site, involving two finishing barns housing up to 1000 pigs/barn. The site had 6 other barns and 6000 pigs that were not directly involved (8000 pigs on site). Pigs were derived from a sow farm that was PRRSv and mycoplasma pneumonia positive, but stable. Pigs were healthy during the test with no clinical signs of either enteric or systemic disease. The study involved 100 pens with 19 pigs each. Males and females were penned separately for the 28 d assay (from 80 to 145 lbs weight) but the response to SBM dose was similar so the data were combined (Boyd and Rush, 2019).
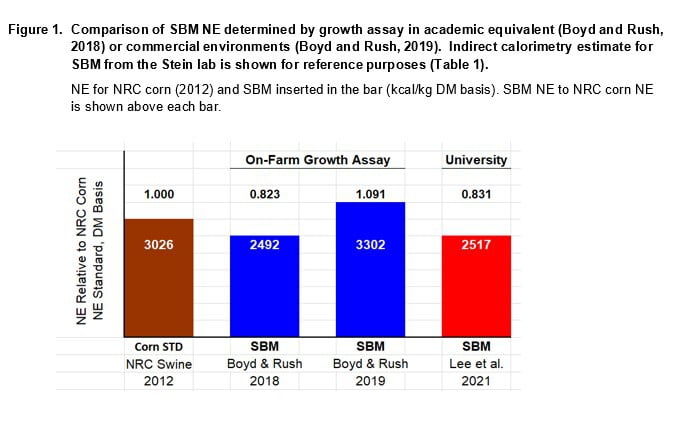
Results for both Hanor assays are shown in Figure 1. Values are expressed on a DM basis and in relation to NRC corn NE (Table 1). Another comparative standard for SBM NE is included in Figure 1. This estimate is from the Stein lab and was determined using indirect calorimetry of pens of pigs fed ad libitum (Table 1).
The first assay with an academic equivalent environment yielded a SBM NE estimate of 2492 kcal/kg DM which compares favorably to the 2502 average for Stein lab studies (Table 1). In apparent contrast, the SBM NE estimate for the commercial environment was 3302 kcal/kg DM. The latter value was greater than NRC corn NE (2012) by a factor of 1.09. In practice, the as-fed feeding value of SBM would be slightly greater because DM content of SBM is often greater than corn at the feed mill.
Results from our first assay confirm SBM NE values from the Stein group, with both being conducted in academic or comparable conditions. The higher SBM NE estimate for the commercial environment (3302 kcal/kg DM) was unexpected; being more than 30% greater than our estimate under a low population-dense environment of the first assay.
This magnitude of difference, combined with the lack of rationale to account for the commercial result, caused us to categorize the commercial result as potentially flawed, so it was indefinitely tabled.
Confirmation that SBM NE is greater in commercial environment
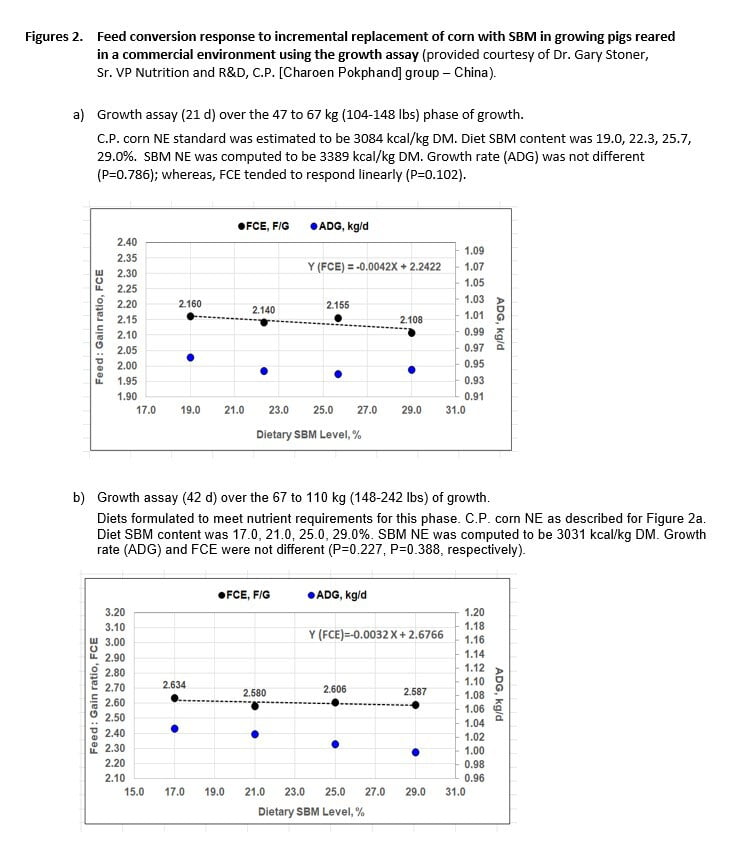
We collaborated with swine nutritionists from the C.P. (Charoen Pokphand) group – China in their study of SBM energetics under commercial conditions (Dr. Gary Stoner, Senior Vice President of Nutrition, Research and Development). The Snyder growth assay was used in two studies (Boyd and Rush, 2017), each involving more than 800 pigs, 4 SBM levels and 18-19 pigs/pen. The first study involved a 21 d test over the 104 to 148 lbs phase of growth, while the second involved a 42 d assay from 148 to 242 lbs body weight. Although pigs during the test did not exhibit clinical signs of disease, health of the sow farm from which they were derived is uncertain.
In both studies, FCE improved as dietary SBM level increased (Figure 2 a,b). SBM NE was estimated to be 3015 kcal/kg as-fed (3389 kcal/kg DM) in the first experiment. In principle, diet FCE could not have improved unless SBM NE was greater than the corn it replaced, and FCE could not have improved at the rate shown (-0.0042 FCE units for each 1.0% SBM added) unless SBM NE was 3015 kcal/kg as-fed. This is equivalent to 1.12 x C.P. China corn NE on as-fed basis (1.10 x C.P. China corn NE, DM basis). The C.P. corn NE was computed from chemical analysis by using the NRC equation 1-8 (2012).
Results for experiment 2 agreed, in principle, with results of the first assay. SBM NE computed to be 2720 kcal/kg as-fed (3031 kcal/kg DM), which is 1.01 x C.P. Group corn NE on an as-fed basis (0.983 x C.P. corn NE, DM basis). Both studies agree with our original finding that SBM NE value is in fact higher for the commercial environment as compared to the academic setting.
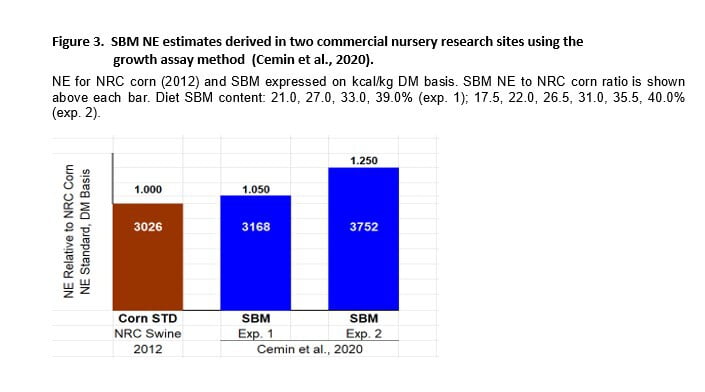
Since our original finding (higher SBM NE) was confirmed by the C.P. China group, other private sector companies have disclosed similar results. Chief among them is the collaboration of Kansas State and JBS Foods – Pork Division that resulted in the peer-review paper by Cemin et al. (2020). Two growth experiments were conducted (21 d each) at 2 commercial research sites and involved more than 6000 nursery pigs (last diet phase). The first study involved 2233 pigs (24 lbs/pig), 92 pens (20-27 pigs/pen) and 4 diets with increasing SBM content. The second study involved 3796 pigs (39 lbs/pig), 84 pens (37-43 pigs/pen) and 6 SBM dietary levels.
The FCE response improved as SBM content increased in both studies (Figure 3). Values were reported on an as-fed basis but corresponding values on a DM basis were 3168 and 3752 kcal NE/kg for experiments 1 and 2, respectively. This equates to 1.05 and 1.25 x NRC corn NE (3026) for nursery pigs in the commercial environment. We note that the 2 studies were conducted in different sites. Their average estimate for SBM NE was 3460 kcal/kg DM (1.15 x NRC corn NE).
Results from this study were surprising in that SBM NE value was high in the nursery phase when pigs were transitioning to higher levels of SBM and the ability of pigs to thrive improved as SBM content increased (data not shown).
An important conclusion from the commercial studies is that results were repeatable; 5 studies in 4 locations by 3 companies agreed with remarkably similar estimates. Further, these results have been replicated by other companies with commercial research facilities (confirmed by confidential personal communication).
How much higher is SBM NE in the commercial environment?
Two important conclusions are apparent:
Stein lab estimates of SBM NE (2502 kcal/kg DM) agree with the Hanor growth assay estimate that was determined in an academic equivalent setting (2492 kcal/kg DM). We consider Stein lab values to be a reliable standard of comparison against commercial environment estimates.\2. SBM NE estimates from the commercial setting are about 30% higher than for the academic environment.
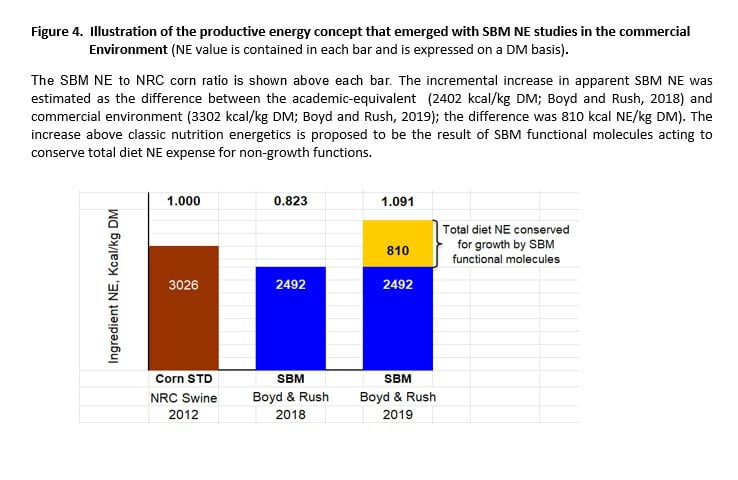
We estimated an incremental increase in SBM NE (810 kcal/kg DM) based on the difference in Hanor growth assay values for the two environments (Figure 4).
Disparity in SBM NE estimates between environmental settings does not imply error for either methodology because the variance is potentially explainable. Stein lab values represent classic substrate energetics, specific to SBM. Commercial SBM NE estimates are also founded on nutritional energetics, but the marginal increase in ‘apparent’ NE may reflect the added influence of functional molecules to conserve total dietary NE for growth, instead of competing processes. The latter is not classic NE, but could represent a metabolic benefit to total diet NE use.
SBM NE values that we presented are summarized according to the environment in which they were determined in Table 2. This information is the basis for our conclusion that apparent NE value for SBM is significantly higher in the commercial setting than for the academic or equivalent environment. The growth assay method confirmed Stein lab estimates for SBM NE where pathogen stress was not a complicating factor (Boyd and Rush, 2018) and was the means for detecting SBM influence on how effectively total diet NE is used for growth (Boyd et al., 2019; Cemin et al., 2020). This is an important compliment to classic energetics in describing the true feeding value of SBM.
Proposed concept for elevated SBM NE value in practice
We propose that the incremental increase in SBM ‘NE’ that we observe in commercial settings is the combination of classic ingredient energetics and possibly an indirect metabolic effect of health-promoting molecules to conserve total diet NE for growth. This is an exciting proposition for animal nutrition which supports the observation that SBM acts in some manner to counter pathogen (or other stressors) related diversion of NE from growth (Boyd et al., 2010). This concept was quantitatively illustrated in Figure 4.
Validation of SBM NE at the commercial level by the Snyder growth assay method led us to expand our understanding of ingredient energetics to the integration of classic energetics with total diet NE conservation by some type of regulation (metabolic, immunological). In other words –
Commercial SBM NE = classic energetics NE + conservation of diet NE for growth
SBM is in a small class of nutraceutical containing feed ingredients that may deliver on the second term in this ‘equation’. SBM is an abundant source of functional molecules that promote health (Moughan and Rutherford-Markwick, 2013). We hypothesize that these molecules counter (in some manner) the need for mounting an immune response to pathogen stressors, even in the subclinical realm. Immune function normally takes priority over growth (survival over production) and imposes a significant energy cost, which varies depending on the degree of pathogen stress. The diversion of NE to counter immune stress occurs in an iterative manner that begins with inflammation and evolves to expansion of immune cell populations and function, each having a significant energy cost (Klasing, 1998; Demas et al., 2011). We recognize that a non-pathogen stressor might also be involved.
Productive energy concept specific to SBM
In view of the incremental increase in SBM NE for the commercial setting that cannot be accounted for by classic energetic measures, we proposed the term ‘productive energy’ (PE) (Boyd and Rush, 2019). This term is specifically applied to SBM to define a formulation value composed of both (1) the historic NE determination, plus (2) amount of total diet NE that was prevented from being diverted to processes other than growth (Figure 4). This may be applicable to other monogastric species (e.g. poultry).
We assert that PE does not interfere with the historic basis for NE, but it quantitatively accounts for the probable metabolic or physiological effect of SBM on diet NE conservation for a productive purpose – growth. This increment is ‘significant’ (approx. 30%) for SBM in the commercial setting and must be taken into account when formulating diets.
In nutrition science, we could define the classic NE value for SBM equivalent to Stein lab reports (e.g. 2502 kcal/kg DM, 47.5% CP), but formulate commercial diets based on PE value that, for example, is equivalent to or 10% greater than corn NE (e.g. 3026 to 3328 kcal/kg DM for NRC 2012 corn). This application appears to be appropriate for growing pigs from late nursery to market.This paper provides another example of the value of combining detailed study at the university level with the scale and environment of the commercial sector in discovery of new opportunities. We believe that this is the first example in animal nutrition where a feed ingredient, with an abundance of functional molecules ‘on-board,’ has proven to increase the total dietary NE partition for growth.
Key Conclusions
SBM NE in commercial environment is equal to or 10% greater than 2012 NRC corn NE value.
Commercial formulations may value SBM NE from 3026 to 3328 kcal/kg DM if related to NRC corn.
A reliable SBM NE value for academic or low population dense settings is 2502 kcal/kg DM.
Functional molecules in SBM may counter pathogen or non-pathogen diversion of dietary NE from growth.
Auxiliary growth assay method backgrounder:Principle and essential elements of growth assay method for validating ingredient NE
The growth assay method of verifying ingredient net energy (NE) in growing pigs has provided values similar to the NRC (2012) for ingredients such as corn germ meal, corn DDGS, choice white grease and wheat midds (similar composition) in the commercial setting. Ingredient NE validation is equally important to studies with the amino acid pattern. Growth assays involve populations, gender, phase of growth and pigs fed ad libitum under varied environmental stresses (e.g. pathogen, crowding) inherent in commercial systems.
The assay is based on comparing the gain to feed response (FCE) for a test ingredient (T-ING) that is substituted for corn (NE reference). No attempt is made to make diets isocaloric since the T-ING NE is unknown. A corn–SBM (C-S) diet with ‘known’ NE content and resulting FCE establishes caloric gain efficiency (mcal NE/kg gain) which is core to computing ING NE. FCE is a discriminating basis for detecting diet energy change with corn substitution. FCE deviance from the C-S standard infers how the T-ING NE compares with the corn that it replaced. The assumption is that the ratio of body protein to lipid deposition is not discernably changed by the T-ING over the test period (28-35 d). We expect this assumption to be true provided that (1) amino acid (6-7 most limiting) and other nutrient needs are met, and (2) growth rate is equivalent among diets. Intake may adjust to meet the NE intake required for tissue growth and composition. NE for the T-ING is computed as the diet NE concentration required to deliver the observed FCE.
The core element of the assay is that corn NE must be accurately predicted since T-ING NE is being quantitatively compared to this standard. Corn is extensively sampled and analyzed for components required by prediction equation 1-8 of the NRC (2012). Components are listed below with auxiliary in vitro measures to verify starch and protein digestibility. In the case of SBM NE validation, a basal SBM level is used for the C-S reference which increases to establish 4-6 diets of the SBM dose-response (Cemin et al., 2020). Crystalline amino acids are adjusted to exceed SID minimums, with other ingredients held constant across diets to limit energy variation to corn displacement; possible exception of meeting STTP specification.(Continued on next page)
Indispensable elements from Snyder growth assay template (Boyd and Rush, 2017):
Corn analyzed for particle size (625-650 u protocol target), DM, CP, C. Fat ether extract, starch, ADF, amino acids. In vitro glucose release by amylase (>65%), protein solubility (>40%) or reactive lysine.
Compute reference corn NE from chemical analysis using NRC (2012) corn DE and equation 1-8. Stein lab corn DE (Table 1) averages 4.4% less than NRC DE.
Test ingredient analyzed; for SBM assay validation, extensive analysis to characterize SBM used, as described in Stein lab papers.
Confirm assay diets by chemical analysis; esp. amino acids, phosphorus for SBM assay.
Feed intake may be slightly altered to meet caloric need without affecting growth rate, however, assay involving high NDF ingredients (e.g. corn germ meal) suppress intake (and growth) at some point which results in inaccurate test ingredient NE. Limit dose-response to the range where growth is not affected.
Regress FCE data to derive regressed (best fit) FCE estimate for each test ingredient dose.
Compute caloric efficiency (mcal NE/kg gain) contributed by ‘known’ diet ingredients using the regressed FCE estimate for each diet. This is the basis for computing the test ingredient NE
Compute NE value required for the test ingredient to deliver the regressed caloric efficiency.
References
Boyd, R.D., M. Sifri, D. Holzgraefe, B. Borg and M. Pope. 2023. Amino acid levels and energy specifications in SBM for poultry and pigs. Feedstuffs, June digital edition, page 10, https://informamarkets.turtl.co/story/feedstuffs-june-2023/page/10.
Lee, S.A., D.A. Rodriguez and H.H. Stein. 2022. Net energy in U.S. soybean meal fed to group-housed growing pigs is greater than calculated book values. 15th Int. Symp. Diges. Physio. Pigs. Anim., Sci. proc. 13 (Issue 2):178.
Boyd, R.D. and C. Rush. 2019. Estimation of soybean meal NE for for healthy growing pigs in the commercial environment by the Snyder feed efficiency growth assay. Hanor Memo. 2019-00. Access via Research Gate, R. Dean Boyd under Technical Reports
Boyd, R.D., M.E. Johnston and C. Zier-Rush. 2010. Soybean meal level modulates the adverse effect of high immune stress on growth and feed efficiency in growing pigs. Proc. 71st Minn. Nutrition Conf. pp. 167-174. U. Minn., St. Paul MN. https://hdl.handle.net/11299/204236
Moughan, P.J. and K. Rutherford-Markwick. 2013. Food bioactive proteins and peptides: antimicrobial, immunomodulatory and anti-inflammatory effects. In Diet, immunity and inflammation. P.C. Calder and P. Yaqoob (ed). Woodland Pub. Limited, 80 High Street, Sawston, Cambridge UK.
Sotak-Peper, K.M., J.C. Gonzalez-Vega and H.H. Stein. 2015. Concentrations of DE, ME and NE in soybean meal produced in different areas of the United States and fed to pigs. J. Anim. Sci. 93:5694-5701. https://doi.org/10.2527/jas2015-9281
Rodriquez, D.A., S.A. Lee and H.H. Stein. 2020. Digestibility of amino acids and concentrations of ME and NE are greater in high-shear dry soybean expellers than in soybean meal when fed to growing pigs. J. Anim. Sci. 98 (7):1-8. https://doi.org/10.1093/jas/skaa215
Lee, S.A., D.A. Rodriquez and H.H. Stein. 2021. Determination of the net energy in soybean meal fed to pigs using indirect calorimetry. Report to United Soybean Board, USB Res. Proj. 2030-353-0515-G
National Research Council (NRC). 2012. Nutrient requirements of swine 11th revised edition. National Academy Press, Washington, DC, USA. https://doi.org/10.17226/13298
Rostagno, H.S. et al., 2017. Brazilian tables for poultry and swine: Composition of feedstuff and nutritional requirements. 4th ed. Dep. Zootecnia, Univ. Fed. Vicosa MG
Boyd, R.D. and C. Zier-Rush. 2017. Practical method for calibrating ingredient NE estimates for growing pigs: Growth assay calibration of wheat midds. Hanor Tech. Memo. H 2008-10. January 2008, DOI:10.13140/RG.2.2.24483.25125
Boyd, R.D. and C. Rush. 2018. Estimation of soybean meal NE for healthy growing pigs in academic-equivalent environment by the Snyder feed efficiency growth assay. Hanor Memo. 2018-00. Access via Research Gate, R. Dean Boyd under Technical Reports
Cemin, H. S., H. E. Williams, M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. Derouchey, K. F. Coble, B. A. Carrender, and M. J. Gerhart. 2020. Estimate of the energy value of soybean meal relative to corn based on growth performance of nursery pigs. J. Anim. Sci. Biotech 11:70. https://doi.org/10.1186/s40104-020-00474-x
Klasing, K.C. 1998. Nutritional modulation of resistance to infectious diseases. Poul. Sci. 77:1119-1125. https://doi.org/10.1093/ps/77.8.1119
Demas, G., T. Greives, E. Chester and S. French. 2011. The energetics of immunity. In Ecoimmunology. G. Demas and R. Nelson. Chap. 8. Oxford Univ. Press. Univ. Oxford, England.
R. Dean Boyd, Ph D. is an adjunct professor of Animal Nutrition at North Carolina State U. and Iowa State U. Aaron M. Gaines, Ph D. is an animal nutritionist and co-founder and managing partner of Ani-Tek Group LLC.