Impacts on B12 and folate concentrations, interrelationships in maternal, fetal fluids
By Jessica G. Syring, Matthew S. Crouse, Tammi L. Neville, Alison K. Ward, Carl R. Dahlen, Lawrence P. Reynolds, Pawel P. Borowicz, Kyle J. McLean, Bryan W. Neville and Joel S. Caton
Limited nutrition for cattle during gestation can arise during times of drought or poor forage quality which can impact fetal programming and development.1 The degree of impact depends on many factors including timing, length and severity of the restriction.
Fetal programming is a process that can have short- and long-term effects on the offspring and is influenced by the maternal environment, including maternal diet and nutrition.2 Fetal programming can occur through a process known as epigenetic modification. Epigenetic modifications include the addition of methyl groups to DNA strands or proteins to alter how genes are expressed. This addition allows gene expression to be changed without changing the actual DNA sequence.
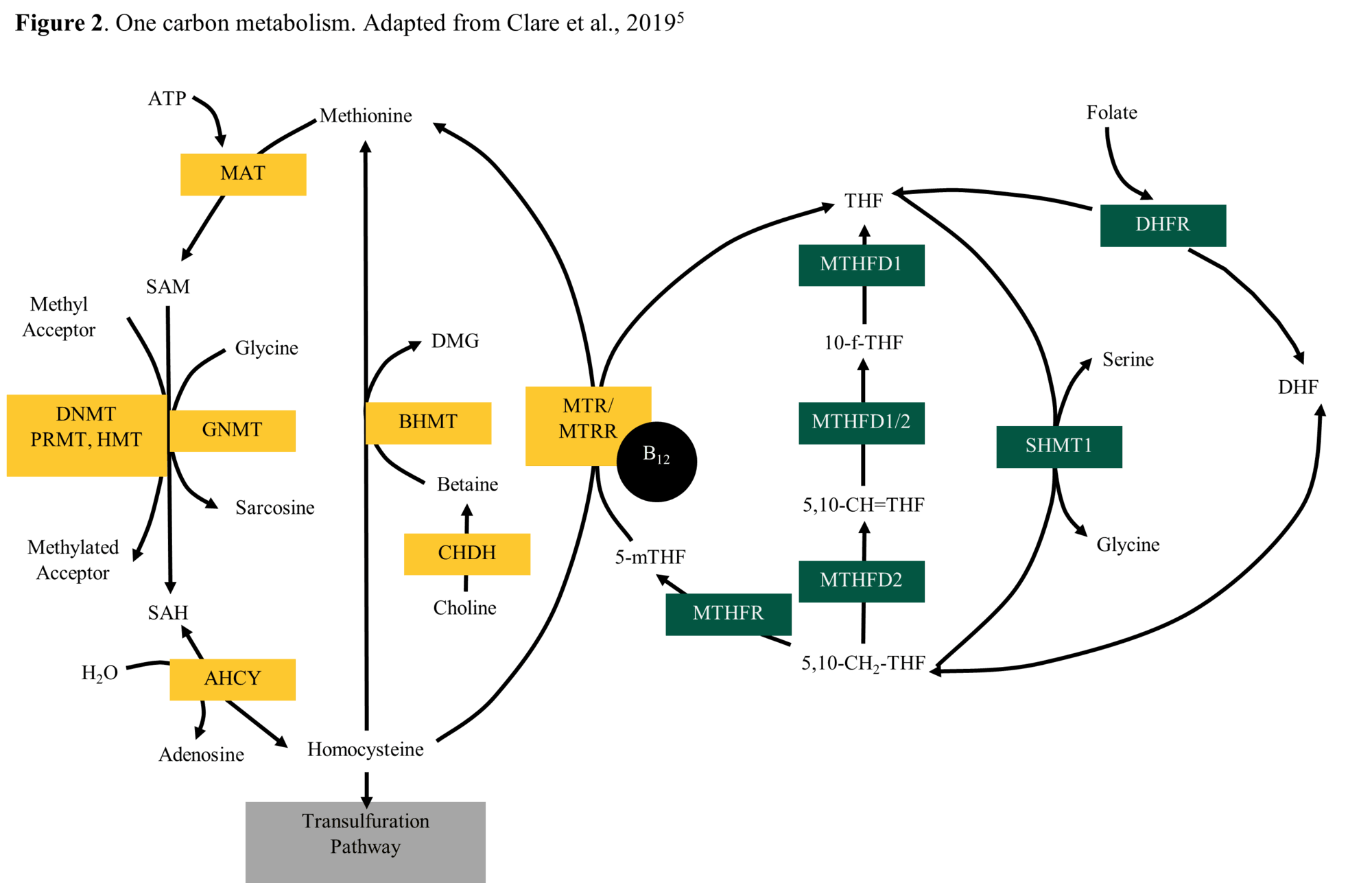
One carbon metabolism is a metabolic pathway involved in DNA and protein methylation. The nutrients involved in one carbon metabolism are called one carbon metabolites (OCM) and include the vitamins folate, choline and B12, as well as the amino acid methionine. By understanding how restricted maternal nutrition affects the abundance of these intermediates, we can gain insight into the effects on the developing fetus.
We hypothesized that moderately restricting maternal nutrition during early gestation would affect OCM concentrations in maternal serum, allantoic fluid (ALF) and amniotic fluid (AMF) and their interrelationships. The objectives of this study were to (1) determine the effects of restricted maternal nutrition from day 0 to 50 of gestation on the concentrations of B12 and folate in maternal serum, ALF and AMF and (2) explore how the concentrations of OCM correlate to each other.
Experimental design and treatments This study used 43 angus-cross heifers that were artificially inseminated using a single sire. Heifers were started on treatment after insemination (day 0). Heifers were fed individually, and treatments were either a control (CON) or restricted (RES) diet.
Heifers on a control diet were fed to gain 1 pound per day and meet 100% of requirements for growth, while heifers on a restricted diet were fed to maintain body weight. The diet consisted of a total mixed ration including grass hay, corn silage, alfalfa haylage, grain and mineral mix, and dried distiller’s grain with solubles to meet protein requirements.
Sample collections Heifers were ovariohysterectomized on day 16, 34 or 50 of gestation. Maternal serum was collected on all three days, but ALF and AMF samples were only collected on day 34 or 50 due to the small size of the fetus and its fluids on day 16 of gestation.
Blood was collected from the maternal jugular vein, allowed to sit at room temperature for 20 minutes to clot, and then centrifuged. The serum was then separated from the sample and stored at -20˚C. Samples of ALF and AMF were collected from the chorioallantois and amnion, respectively, snap frozen, and also stored at -20˚C. For B12 and folate analysis, samples were sent to IDEXX BioAnalytics (N. Grafton, MA).
Other OCM (methionine, homocysteine, serine and glycine) presented in this paper were previously analyzed by and are presented in a paper by Crouse et al. (2019).3 Briefly, serum and fetal fluid sample amino acid concentrations were analyzed using ultra-performance liquid chromatography (UPLC).3
Statistical analysis For B12 and folate analysis, the GLM procedure of SAS (SAS Inst. Inc.) was used. Individual heifer served as the experimental unit, and treatment, day of gestation and their interaction was included in the first model.
The second model also had heifer as the experimental unit, and contained treatment, fluid type and their interaction. If no significant interactions (P > 0.05) were found, main effects were reported. Means were separated using LSMEANS. Values were considered significant at P ≤ 0.05.
Pearson correlation coefficients for OCM were calculated using PROC CORR. A negative correlation is indicated by a negative ¬r-value, and a positive correlation is indicated by a positive r-value. Correlation coefficients were considered significant with a P-value ≤ 0.05.
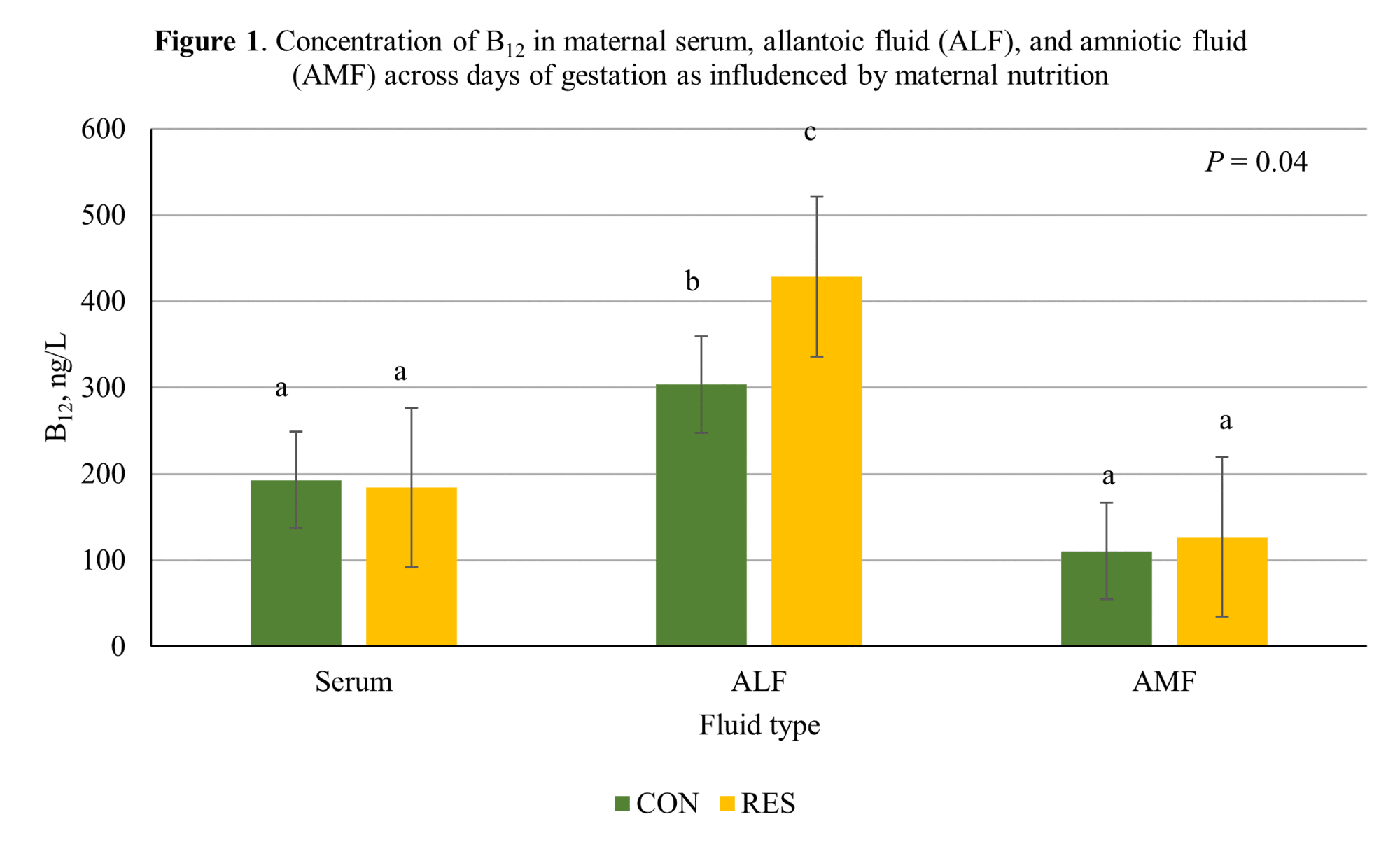
Results and discussion Results for B¬12 and folate analysis can be found in Tables 1 and 2. The restricted treatment increased B12 and folate concentrations (P = 0.04 and P = 0.03, respectively) in ALF. Additionally, folate concentrations were higher in day 50 fetuses compared to day 34 fetuses (P = 0.003).
Two main functions of the allantois are gas exchange and fetal waste storage, with allantoic fluid found within it. Concentrations of B12 were greatest in RES ALF, intermediate in CON ALF, and least in both CON and RES serum and CON and RES AMF (P < 0.04). This difference is illustrated in Figure 1.
Finally, concentrations of folate were greatest in ALF, intermediate in maternal serum and least in AMF (P < 0.001). These fluid differences may not be surprising considering they each have different functions and make ups.
Serum is the liquid part of blood that does not contain any clotting factors, red or white blood cells or platelets. It helps in the transportation of substances throughout the bloodstream.
As mentioned previously, ALF plays a role in fetal waste collection and gas exchange, although some nutrient absorption is also thought to take place as well. It is composed of water, nutrients and hormones.
Finally, AMF starts off mostly as water, but changes composition as the fetus matures, and functions to protect the developing fetus.
Correlations between OCM can be found in Table 3. As serum B12 concentrations changed, serum methionine concentrations changed in the same direction (r = 0.48). This relationship is not surprising since B12 is needed to remethylate homocysteine to methionine (Figure 2).
However, when serum folate concentrations changed, AMF homocysteine concentrations changed in the opposite direction (r = -0.64). Allantoic folate and AMF glycine concentrations changed in the same way (r = 0.66).
Finally, AMF folate concentrations changed in the same way as ALF methionine (r = 0.48), ALF serine (r = 0.60), and AMF serine (r = 0.62). Folate drives methionine synthase reactions, which reduces homocysteine concentrations and increases methionine concentrations (Figure 2).
Crouse et al. (2019) found an increase in serum serine in CON heifers compared to RES heifers and an increase in ALF glycine in day 50 fetuses compared to day 34.3
They also found higher concentrations of AMF methionine, serine and glycine in day 34 fetuses compared to day 50.3 Amino acids act as building blocks for other biological molecules, as well as antioxidants and pathway signalers. Thus, they are important factors as well in fetal development.
Kwon et al. (2003) also observed alterations in amino acid concentrations in ovine maternal and fetal fluids.4 Changes in ALF concentrations were thought to be a result of regulating electrolyte concentrations of ALF, while changes in AMF amino acid concentrations were a result in the metabolism and production of amino acids by the fetus.4
Because metabolic pathways are interconnected, changes in one metabolite results in the change of other metabolites. Specifically, in one carbon metabolism, folate is needed for the production of the amino acids serine, glycine and methionine.
By reducing folate concentrations, the production of those amino acids is reduced. The amino acid homocysteine is remethylated to produce methionine. Thus, if folate supply is reduced, the amount of methionine produced is reduced, and the levels of homocysteine are increased. A different pathway, transsulfuration, uses the excess homocysteine, increasing its metabolic products.
Implications The results of this study show that limited maternal nutrition increases B12 and folate concentrations in fetal allantoic fluid, and those changes correlate to changes in other metabolites in one carbon metabolism. These changes may be brought about by possible alterations in maternal metabolic pathways in response to global nutrient restriction. Metabolic pathways are interconnected, so a change in one can lead to a change in another.
What impacts these maternal shifts have on fetal development have yet to be studied in depth, but these findings may suggest that the increase in concentrations of B12 and folate in fetal fluids means less of these nutrients for fetal growth and development is used, as fetal growth is often reduced when maternal nutrition is restricted. These findings also give us insight towards developing strategic supplementation plans during early pregnancy.
References 1Wu, G., F. W. Bazer, T. A. Cudd, C. J. Meininger, and T. E. Spencer. 2004. Maternal nutrition and fetal development. J. Nutr. 134:2169–2172.
2Caton, J. S., M. S. Crouse, K. J. McLean, C. R. Dahlen, A. K. Ward, R. A. Cushman, A. T. Grazul-Bilska, B. W. Neville, P. P. Borowicz, and L. P. Reynolds. 2020. Maternal periconceptual nutrition, early pregnancy, and developmental outcomes in beef cattle. J. Anim. Sci. 98:1–16.
3Crouse, M. S., N. P. Greseth, K. J. McLean, M. R. Crosswhite, N. N. Pereira, A. K. Ward, L. P. Reynolds, C. R. Dahlen, B. W. Neville, P. P. Borowicz, and J. S. Caton. 2019. Maternal nutrition and stage of early pregnancy in beef heifers: impacts on hexose and AA concentrations in maternal and fetal fluids. J. Anim. Sci. 97:1296–1316.
4Kwon, H., T. E. Spencer, F. W. Bazer, and G. Wu. 2003. Developmental changes of amino acids in ovine fetal fluids. Biol. Reprod. 68:1813–1820.
5Clare, C. E., A. H. Brassington, W. Y. Kwong, and K. D. Sinclair. 2019. One-carbon metabolism: linking nutritional biochemistry to epigenetic programming of long-term development. Annu. Rev. Anim. Biosci. 7:263-287.
Mention of a trade name, proprietary product, or specific agreement does not constitute a guarantee or warranty by the USDA and does not imply approval to the inclusion of other products that may be suitable. USDA is an equal opportunity provider and employer.
Means3
CON
RES
d16
d34
d50
SEM4
Trt
P-values
Day
Trt x Day
B12, ng/L
Folate, µg/L
193.3
6.9
184.0
7.3
196.0
7.5
195.3
6.7
175.6
--
33.50
0.65
0.67
0.47
0.66
0.18
0.83
298.9
8.9
423.0
12.2
353.4
13.7
368.6
7.4
61.10
1.62
0.04
0.03
0.79
0.003
0.09
0.07
110.4
1.7
126.6
1.4
14.60
0.16
0.43
0.10
1Day of gestation (Day) = days after insemination. 2ALF = allantoic fluid; AMF = amniotic fluid. 3Treatment (Trt): Control (CON), restricted (RES); day of gestation: d16, d34, d50. 4Greatest SEM for the treatment × day interaction.
Fluid1 Item2
Serum
ALF
AMF
Treatment Mean3
P-values Treatment
Fluid
Treatment x Fluid
CON RES Fluid5
303.3b 428.3c 365.8
110.4a 126.6a 118.5
202.1 246.3
Table 2. Concentrations of vitamin B12 and folate in maternal serum, allantoic fluid and amniotic fluid across days 16, 34 and 50 of gestation as influenced by treatment.
1ALF = allantoic fluid; AMF = amniotic fluid. 2CON = heifers fed a diet that met 100% of NRC requirements to gain 0.45 kg daily; RES = heifers restricted to 60 percent of the CON diet. 3Mean B12 and folate concentrations of treatment groups across fluids. 4Greatest SEM for the treatment × fluid interaction (Allantoic CON n = 13, Amniotic CON n = 5, Serum CON n = 17, Allantoic RES n = 16, Amniotic RES n = 6, and Serum RES n = 23) for B12; average SEM for the day × treatment interaction (Allantoic CON n = 13, Amniotic CON n = 5, Serum CON n = 12, Allantoic RES n = 16, Amniotic RES n = 6, and Serum RES n = 16) for folate. 5Mean vitamin B12 or folate concentration across treatments within fluid. a-cMeans without a common superscript differ (P < 0.05) for treatment × fluid. d-fMeans within a row without a common superscript differ (P < 0.05) for main effect of fluid.
B12 Serum
Folate Serum
Table 3. Pearson correlation co-efficents for relationship between B12 and folate concentrations (in maternal serum, allantoic fluid and amniotic fluid) and methionine, homocysteine, serine and glycine concentrations in maternal serum, allantoic fluid and amniotic fluid1.
1ALF = allantoic fluid; AMF = amniotic fluid. *P < 0.05 **P < 0.01 †0.1 > P < 0.05.
Syring, Tammi Neville, Ward, Dahlen, Reynold, Borowicz and Caton are with the Center for Nutrition and Pregnancy, North Dakota State University; Crouse and Bryan Neville are with the USDA, ARS, U.S. Meat Animal Research Center; and McLean is with the Department of Animal Science, University of Tennessee.