It impacts more than just the cow!
By Hannah Speer, Harvey Freetly, Karla Wilke and Mary Drewnoski
Vitamin A has several important roles in the body. It is well-known for its role in vision, but it is also important for proper immune function and epithelial integrity, specifically in the gastrointestinal and respiratory tracts1. Clinical deficiency is unlikely to occur in most cases, but marginal deficiencies can still impact cow productivity and potentially calf health.
Calves are born with very low vitamin A stores, and their primary source at birth is colostrum. Vitamin A concentrations in colostrum have been reported to be six2 to 143 times greater than that of milk, so colostrum is critical for establishing vitamin A stores in the young calf. Calves that do not get enough vitamin A from colostrum can be at increased risk for diarrhea and respiratory disease.
Fresh green forage contains high amounts of beta carotene, a precursor that can be used by the cow to synthesize the vitamin A she needs to support a variety of biological functions. Any excess vitamin A that is made can be stored in the liver and used during times when dietary vitamin A intake is low.
Conversely, cows fed diets consisting primarily of stored forages and concentrates may be at risk for vitamin A deficiency because these feedstuffs are low in beta carotene. Low vitamin A levels in the cow’s diet during late gestation could potentially impact vitamin A concentrations in colostrum, which may lead to a calf becoming deficient and subsequently impact calf health.
No research has been conducted in beef cows to know what amount of supplemental vitamin A is needed to ensure both cows and their calves achieve adequate vitamin A status. The objectives of these studies were to identify the relationship between cow and calf vitamin A status using plasma and liver samples, and to understand the effect of supplemental vitamin A level from mid-gestation to early lactation on liver vitamin A concentrations in the cow and subsequently her calf.
Our hypothesis was that cows having liver vitamin A concentrations that fell within adequate reference ranges would result in her calf having adequate liver vitamin A concentrations. Additionally, it was also hypothesized that current NASEM recommendations for supplemental vitamin A would not be sufficient for gestating cows that have been fed stored forages and concentrates for an extended period.
Experiment 1The study was conducted at the U.S. Meat Animal Research Center near Clay Center, Nebraska. Multiparous beef cows that had previously been grazing on pasture (6.4 ± 1.2 years of age; n = 120) in mid-gestation were assigned to receive 9,638 IU/d vitamin A (n = 30) or 24,973 IU/d vitamin A (n = 90). These levels were approximately one-third and two-thirds of the current NASEM recommendation of 2,800 IU/kg DM (33,000 IU/d in this study) for gestating beef cows weighing 1,300 lbs consuming 2.0% of body weight in DM per day.
Cows were individually supplemented in Calan gates from 111 days pre-calving to 32 days post-calving. Their diet consisted of alfalfa hay, corn silage and a supplemental pellet that contained the vitamin A, which was provided as retinyl acetate. Basal diet vitamin A concentration was calculated to be 490 IU/kg DM based on its beta carotene content, so mean vitamin A intake from the basal diet was 4,583 ± 649 IU/d.
For assessing vitamin A status, liver biopsies and blood samples were collected at day 0 (111 days pre-calving) and day 144 (32 days post-calving), and calves were sampled at 32 ± 7 days of age. Vitamin A concentrations, measured as retinol, were analyzed in plasma and liver, and Pearson correlations were used to test for linear relationships between cow liver and plasma retinol concentrations, calf liver and plasma retinol concentrations, and liver retinol concentrations between the cow and her calf.
Experiment 2This study took place at the Panhandle Research and Extension Center in Scottsbluff, Nebraska. Multiparous beef cows (n = 54) that had been fed in the drylot for a year or more were stratified by body condition score and source and assigned to a pen.
Pens (n = 3 per treatment) were then randomly assigned to receive 1 of 3 supplemental vitamin A levels: the current NASEM recommendation for gestating beef cows (31,000 IU/d; 1X), 3 times (93,000 IU/d; 3X), or 5 times the current NASEM recommendation (155,000 IU/d; 5X). The 1X level was selected in this study assuming a cow weight of 1,200 lbs that consumed 2.0% of body weight in DM per day.
Prior to treatment initiation, all cows were receiving the 1X supplemental level. Treatments were initiated in mid-gestation and concluded 32 days post-calving. Cows were limit-fed a diet consisting of wheat straw, corn silage, and wet distillers grains. Vitamin A, as retinyl acetate, was added to the diet via a micronutrient machine.
Liver biopsies were collected for retinol analysis on cows 24 days before treatment initiation, d 40 and d 81 of supplementation, and both cows and calves were sampled 32 d post-calving (165 ± 22 d SD of supplementation).
Results Experiment 1Because cows had recently spent time on green grass, initial liver retinol concentrations (mean 830 µg/g DM) of cows were well above adequate. By 32 days post-calving, mean cow liver retinol concentration (482 ± 182 SD µg/g DM) had decreased but was still considered adequate based on the current reference range of 300–700 µg/g DM.
Cow plasma retinol (272 ± 40 SD ng/mL) was slightly below the reference range of 300–800 ng/mL. No linear relationship (P = 0.10; r = 0.16) was observed between liver and plasma retinol in cows, which is not surprising because plasma retinol concentrations are homeostatically regulated and will not fluctuate unless liver vitamin A concentrations are very low4.
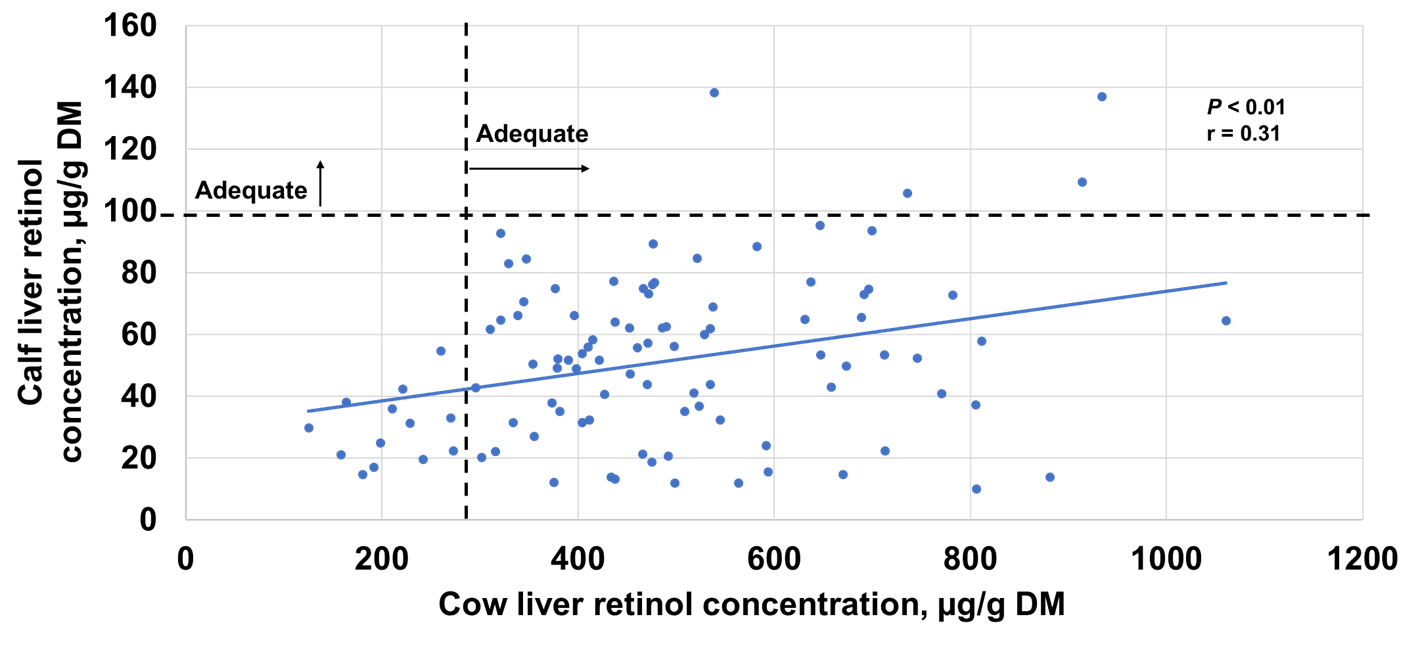
A positive correlation (P < 0.01; r = 0.37) was detected between calf liver (51 ± 27 SD µg/g DM) and plasma (190 ± 47 ng/mL) retinol concentrations. Both were below what would be considered adequate (100–350 µg/g DM in liver; 225–325 ng/mL in plasma) for calves at 32 days of age. It is suspected that a correlation was observed here because the majority of calves had liver retinol concentrations less than 100 µg/g DM (Figure 1), which may have caused fluctuations in plasma retinol concentrations.
There was a positive correlation (P < 0.01; r = 0.31) between cow and calf liver retinol (Figure 1), suggesting that as cow retinol liver concentrations increased, calf liver retinol concentrations increased. However, it appears that despite cows having adequate liver retinol concentrations, when supplemental vitamin A levels below NASEM recommendations were fed, it did not result in calf liver retinol stores that would be considered adequate given current reference ranges. This is likely because cow liver retinol stores are not the only contributor to vitamin A in colostrum.
Research in beef cattle indicates cow stores only contribute about 40% of the vitamin A found in colostrum, while the other 60% comes from the cow’s diet3,5. Therefore, dietary vitamin A levels the cow receives during late gestation, as well as her liver vitamin A stores, affect the amount of vitamin A her calf receives to build its own liver vitamin A stores.
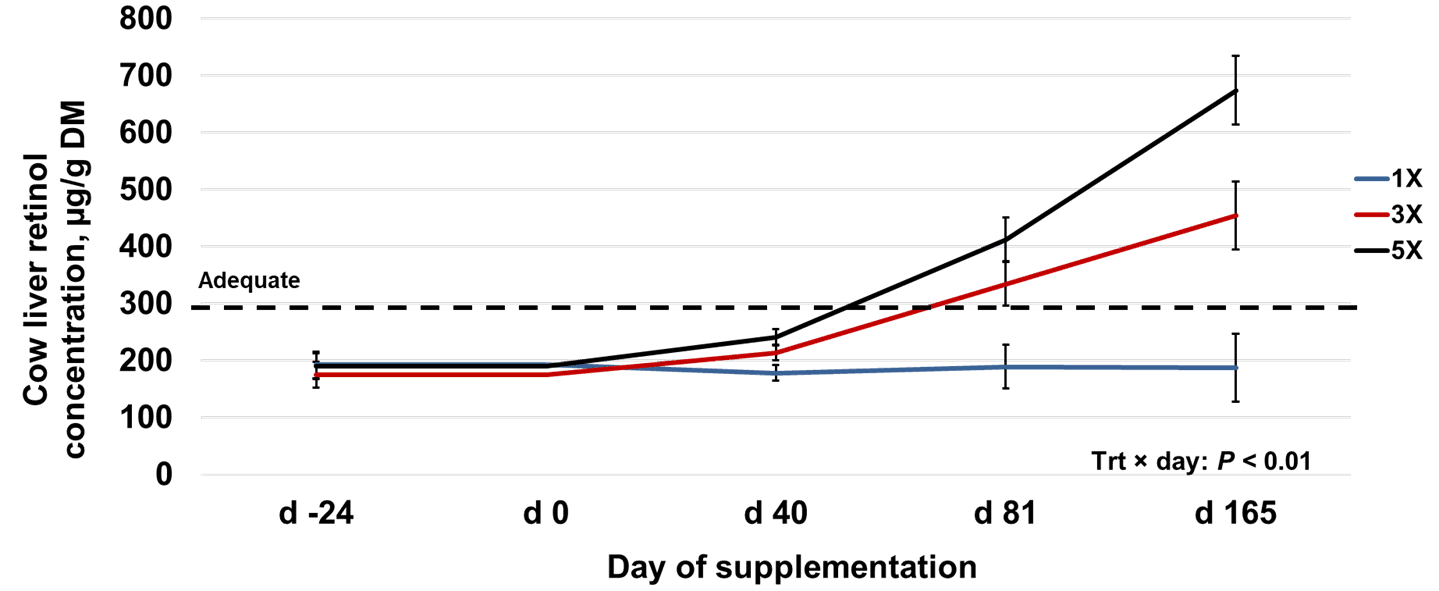
Results Experiment 2No differences (P = 0.86) in initial cow liver retinol concentration (mean 186 µg/g DM; Figure 2) were observed between treatments. Cows were receiving the 1X supplemental vitamin A level before the study, suggesting that the current supplemental vitamin A recommendation of 31,000 IU/d was not enough to get cows to adequate liver retinol concentrations (300–700 µg/g DM).
A significant treatment × day interaction (P < 0.01) was observed for cow liver retinol. On d 40, cows in 1X had liver retinol concentrations (178 µg/g DM) that were not different (P = 0.12) from 3X (213 µg/g DM) but lower (P = 0.02) than 5X (241 µg/g DM), while 3X and 5X did not differ (P = 0.21). Liver retinol on d 81 was lower (P < 0.05) in 1X (189 µg/g DM) compared to 3X (334 µg/g DM) and 5X (412 µg/g DM), which did not differ (P = 0.20).
For cow liver retinol post-calving, 1X (187 µg/g DM) was lower (P < 0.05) than 3X and 5X, and 3X (454 µg/g DM) was lower (P < 0.05) than 5X (674 µg/g DM). Liver retinol concentrations of 1X cows remained below adequate reference ranges (300–700 µg/g of DM) throughout the study, whereas 3X and 5X were elevated into the adequate range by d 81 of supplementation.
Calf liver retinol concentration also differed among treatments (P = 0.01; Figure 3), as calves of cows in 1X had lower (P < 0.05) liver concentrations than 3X and 5X calves which did not differ (P = 0.12). Liver retinol concentrations considered adequate for calves at 32 days of age (100–350 µg/g of DM) were not observed in 1X calves (51 µg/g DM) but were observed in calves from 3X and 5X cows (119 and 165 µg/g DM, respectively).
These results suggest that for cows fed stored feeds long term, supplementing cows with the current NASEM recommendation for vitamin A will not result in their calf’s liver vitamin A concentrations being within the adequate reference range.
Our data also suggests that cows with initially low liver retinol stores needed to be fed 93,000 IU/d (3 times the NASEM recommendation) of vitamin A to achieve adequate liver retinol concentrations. However, this amount did appear to result in continuously increasing liver stores. More research is needed to understand the exact amount of supplemental vitamin A required to maintain cow liver retinol concentrations in the adequate range and ensure adequate concentrations in the colostrum for the calf.
ImplicationsThe calf is at greatest risk of vitamin A deficiency when cow vitamin intake is low. Unlike our initial hypothesis, a cow that has adequate liver vitamin A stores at the time of calving does not ensure that the calf will also have adequate liver vitamin A stores. Both cow liver stores and cow vitamin A intake influence vitamin A levels in colostrum, so it benefits the calf if the cow has both adequate liver vitamin A stores and receives adequate supplemental vitamin A in late gestation. These data suggest that feeding the NASEM supplemental vitamin A recommendations to cows fed stored feeds long term does not result in adequate beef cow or calf liver retinol concentrations.
References1Semba, R. D. 1998. The role of vitamin A and related retinoids in immune function. Nutrition Reviews. 56:S38–S48.
2Schweigert, F. J. 1989. Effect of gestation and lactation on lipoprotein pattern and composition in dairy cows. J. Anim. Physiol. Anim. Nutr. 63:75–83.
3Branstetter, R. F., R. E. Tucker, G. E. Mitchell, Jr., J. A. Boling, and N. W. Bradley. 1973. Vitamin A transfer from cows to calves. Internat. J. Vit. Nutr. Res. 43:142–146.
4Olson, J. A. 1984. Serum levels of vitamin A and carotenoids as reflectors of nutritional status. J. Natl. Cancer Inst. 73:1439–1444.
5Tomlinson, J. E., G. E. Mitchell, Jr., N. W. Bradley, R. E. Tucker, J. A. Boling, and G. T. Schelling. 1974. Transfer of vitamin A from bovine liver to milk. J. Anim. Sci. 39:813–817.
Speer is a graduate research assistant, Wilke is an associate professor and range management cow/calf specialist, and Drewnoski is a beef systems specialist; all with the University of Nebraska-Lincoln. Freetly is a nutrition, growth and physiology research leader with the USDA Agricultural Research Service.